
2019 North Dakota Beef and Sheep Report (AS1938, Sept. 2019)

(Photo by Lauren Hanna, NDSU)
This is the eighth year that I have been the coordinator and editor of the North Dakota Beef Report (the last two years as the North Dakota Beef and Sheep Report) and I still very much enjoy this activity as it is an important means to report our research findings to producers and industry personnel across North Dakota and beyond. It also allows me to reflect about the quality and breadth of beef cattle and sheep research across North Dakota.
The beef and sheep research programs at the North Dakota Agricultural Experiment Station’s Main Station in Fargo and the Research Extension Centers across North Dakota are dedicated to serving the producers and stakeholders in North Dakota by developing new knowledge and technology to improve the management, efficiency and production of high-quality cattle, beef, sheep and lamb. This report includes a broad range of research from on-campus departments, schools and centers, as well as Research Extension Centers across the state, and provides producers and stakeholders with one document that contains reports of beef- and sheep-related research conducted at NDSU each year.
I want to again thank Ellen Crawford and Deb Tanner for their great assistance in editing and formatting the reports so that we can publish a great statewide combined report. Also, thanks to the contributors to the report, and to the staff and students who help will the research, teaching and Extension activities related to beef cattle and sheep.
Finally, thanks to the funders of the grants that help fund the research projects and students/staff working on the projects. We truly appreciate your contributions to our research program. Without this support, the research would not be possible.
If you should have any questions about the research highlighted in this report, please do not hesitate to contact me or any of the authors of the individual reports. Thanks for your encouragement and support of beef cattle and sheep research in North Dakota.

(Photo by Kacie McCarthy, NDSU)
Grazing / Forage Nutrition
Utilizing an electronic feeder and ear-tag accelerometer to measure mineral and energy supplement intake and reproductive behavior in beef heifers grazing native range
Kacie L. McCarthy1, Sarah R. Underdahl1, Michael Undi2, Stephanie Becker2 and Carl R. Dahlen1
1Department of Animal Sciences, NDSU
2Central Grasslands Research Extension Center, NDSU
The objectives of this study were to develop a Mobile Cow Command Center (MCCC) for use to 1) examine the relationship between mineral and energy supplementation on intake and feeding behavior on native range and 2) examine reproductive behavior of heifers on native range utilizing the CowManager system. The MCCC paired two commercially available technologies, a SmartFeed device, which monitors intake, and a CowManager system, which monitors cow reproductive, and feed-related and health-associated data in a single trailer unit that can be transported and utilized anywhere cattle are managed. Our results show that the feed-controlling portion of the MCCC can be used for precision feeding of individuals in expansive group-managed scenarios. Further, the estrus and health alert functions of the CowManager system were found to be unreliable triggers for management responses that could lead producers to inappropriate conclusions about the status of their herds.
Summary
Crossbred Angus yearling heifers (n = 60) at the Central Grasslands Research Extension Center (Streeter, N.D,) were used to evaluate an electronic feeder to measure mineral and energy supplement intake and an ear-tag accelerometer to indicate feeding activity and reproductive behavior in heifers grazing native range. Heifers were fitted with radio frequency identification (RFID) ear tags that allowed access to an electronic feeder from which supplements were delivered. Heifers were assigned randomly to one of three dietary treatments: 1) no access to feed supplements (n = 20); 2) free-choice access to mineral supplement (n = 20) or 3) free-choice access to energy supplement (n = 20). Heifers also were fitted with a CowManager tag that uses sensors to monitor cow reproductive (estrus alerts), feed-related (eating, rumination and activity level) and health-associated (body temperature) data. Heifers were artificially inseminated utilizing sexed semen and turned out to graze at the initiation of the study. Consecutive day weights, along with blood and liver biopsy samples, were taken at the beginning and end of the study. Electronic feeders effectively controlled intake where, by design, heifers in the NRG treatment (44.07 ± 1.28 ounces/day) consumed more (P < 0.001) energy supplement, compared with CON (0.23 ± 1.28 ounces/day) or MIN (0.007 ± 1.28 ounces/day) heifers. Final liver mineral concentrations in NRG heifers were greater (P < 0.05) for selenium (Se), iron (Fe) and cobalt (Co), compared with CON and MIN heifers. At the end of the monitoring period, NRG heifers had greater (P = 0.03) serum glucose concentrations, compared with CON and MIN heifers. The MCCC units were deployed successfully and serve as portable units that use solar power to run individual feeders and upload data to cloud-based data acquisition platforms.
Introduction
As technology is advancing, some sectors of agriculture are implementing new innovations rapidly. However, the beef industry is lagging behind other industries in the rate of adoption.
Several reasons likely exist for this adoption lag, foremost of which are the lack of comprehensive technological solutions that can be implemented in expansive pasture settings, and the lack of solutions from which management decisions can be made during the life of the individual. Each individual in a herd of cattle is unique, with variations in the stage of production, specific nutritional needs and health status, and these variations change throughout the production year.
To maintain targeted production levels beneficial for growth and reproductive performance and offset forage nutritive decline throughout the grazing season (Schauer et al., 2004; Cline et al., 2009), producers often provide mineral and/or protein and energy supplements to grazing cattle. An issue observed with providing supplements on pasture is the large variability in consumption by individuals within a group (Tait and Fisher, 1996; Bowman and Sowell, 1997; Cockwill et al., 2000; Patterson et al., 2013), which is largely unseen and unknown by personnel managing the cattle.
In addition, frequent observation of activity and reproductive behavior of grazing cattle often is difficult due to the expansive area occupied by pastures. However, electronic systems are available to monitor feeding, activity and reproductive-related behavior.
Activities reported herein are aimed at pairing technologies into a single Mobile Cow Command Center that would allow for precision management of individuals within a herd to optimize production efficiency, improve animal health and enhance profitability.
Our objectives were to develop a Mobile Cow Command Center (MCCC) for use in 1) examining the relationship between mineral and energy supplementation on intake, liver mineral concentrations and metabolites in heifers being managed on native range and 2) examining activity and reproductive behavior of heifers on native range.
Experimental Procedures
All animal procedures were conducted in accordance with the Institutional Animal Care and Use Committee at North Dakota State University.
Mobile Cow Command Center Units
Each of two MCCCs were developed by pairing two commercially available technologies into single trailer units that can be transported and function anywhere cattle are managed. The first technology is the SmartFeed device (C-lock Inc., Rapid City, S.D., see www.c-lockinc.com), which is a self-contained system designed to measure supplement intake and feeding behavior from individual cattle in group settings. The system is solar powered and includes a radio-frequency identification (RFID) reader, weigh scales, access control gate, a feed bin and a cloud-based interface that continuously logs feed intake and feeding behavior data.
The second technology in the MCCC was the CowManager system (distributed by Select Sires in the U.S., see www.cowmanager.com/en-us). It uses RFID tags and additional sensors to monitor cow reproductive (estrus alerts), and feeding-related (eating, rumination and activity level) and health-associated (body temperature) data.
Data were received by a router attached to a computer in each MCCC that automatically uploaded the data for viewing on any device with an internet connection. Two SmartFeed units and controlling hardware and the CowManager systems were placed in each of two enclosed trailers with open feed access areas and retractable wheels for easy transport.
Training Period
The MCCC units each were placed into a separate dry lot heifer development pen (n = 63 per pen) at the Central Grasslands Research Extension Center (CGREC) for a two-week period. Corn silage was placed into the feed bins and heifer intake was monitored. Only heifers with a history of feed consumption from the feeders were used for the experiment.
Heifer Selection
All heifers were bred via artificial insemination (AI) to a common sire, with pregnancy detection via transrectal ultrasonography (7.0-MHz transducer, 500 V Aloka, Wallingford, Conn.) 34 days after AI.
Grazing Period
Sixty crossbred yearling Angus heifers were managed as a single pasture group with free access to native range grazing and were assigned randomly to one of three dietary treatments: 1) no access to feed supplements (CON; n = 20), 2) free choice access to mineral supplement (MIN; Purina Wind and Rain Storm [Land O’Lakes Inc.], n = 20) or 3) free choice access to energy supplement (NRG; Purina Accuration Range Supplement [Land O’Lakes Inc.], n = 20). The MIN and NRG supplements were delivered via the MCCC SmartFeed units, and only heifers assigned to the respective treatments were allowed access to the feeders through the web-based controlling interface.
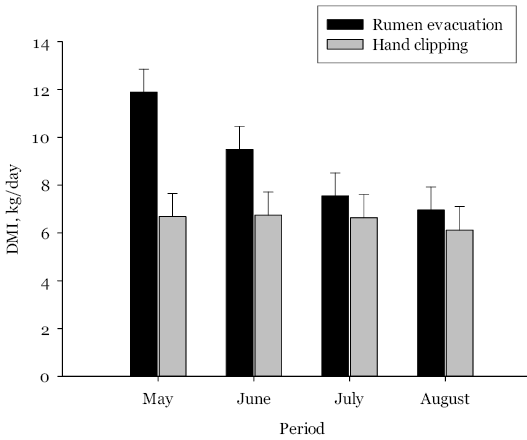
Because few heifers consumed either supplement early in the grazing season (Figure 1), feed intake data were summarized during a 57-day period: from the time of pregnancy diagnosis (July 25) until removal from pasture (Sept. 19, 2019). Heifers assigned to MIN and NRG treatments that did not consume the respective supplements were placed into the CON treatment for analysis resulting in n = 29 for CON, n = 18 for MIN and n = 13 for NRG.
Estrus-related events were generated via the CowManager system and were listed as in heat, potential or suspicious. Heifers were monitored for return to estrus after AI and ultrasound was used to confirm pregnancies.
A retrospective analysis was conducted to determine the accuracy of estrus-related alerts generated via the CowManager system versus a known pregnancy status determined via ultrasound. Similarly, a retrospective analysis was conducted to evaluate the accuracy of health events that were flagged via the CowManager system (reported as sick, very sick or no movement) by comparing with treatment logs generated by the animal care staff at the CGREC. The CowManager system also reported the minutes spent each day in activity categories including eating, ruminating, not active, active and highly active.
Blood samples were collected via jugular venipuncture into serum tubes. Serum samples were analyzed for glucose and NEFA concentration.
Samples of liver were collected on the first and final day of monitoring via biopsy from a subset of heifers from each respective treatment (n = 24; eight per treatment). Liver samples were sent to Michigan State University and were evaluated for concentrations of minerals using inductively coupled plasma mass spectrometry.
Analysis
Data were analyzed as a completely randomized design. All data were analyzed using the GLM procedure of SAS. Data were considered significant at P < 0.05.
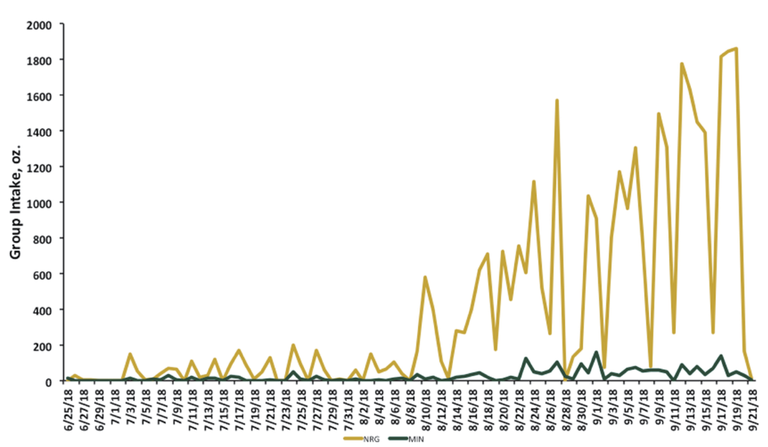
Figure 1. Beef heifer intakes of mineral (MIN) and energy (NRG) supplements during the grazing period.
Results and Discussion
Intake of energy and mineral supplements was very low during the early portion of the grazing season but began to increase in mid-August as the quality of native range declined (Figure 1). From July 25, 2018, until Sept. 19, 2018, heifers in the MIN treatment (1.74 ± 0.12 ounces/day) consumed more (P < 0.001) mineral, compared with heifers in the CON (0.09 ± 0.12 ounce/day) and NRG treatments (0.07 ± 0.12 ounce/day), and heifers in the NRG treatment (44.07 ± 1.28 ounces/day) consumed more (P < 0.001) energy supplement, compared with CON (0.23 ± 1.28 ounces/day) or MIN (0.007 ± 1.28 ounces/day) heifers.
Activity data from the CowManager tags indicate that time spent eating, ruminating, not active or active were not influenced by treatment (P ≥ 0.32; Table 1). However, heifers in the NRG treatment spent 20 more (P = 0.007) minutes on a daily basis being highly active, compared with heifers in the other treatments.
The observed additional time NRG heifers spent being highly active was likely related to competition for energy supplements at feeders, where 13 heifers were competing for two feeding spaces. Interestingly, treatment did not influence weight gain (P = 0.93) during the monitoring period, with heifers gaining an average of 1.01 pounds/day.
Evaluation of estrus data revealed that 16 of 28 heifers confirmed pregnant via ultrasound (57%) were incorrectly identified as displaying some type of estrus behavior (two reported as in heat, 11 reported as potential and three reported as suspicious). Additionally, 146 health alerts were generated, but only 13 heifers needed clinical treatment. An additional nine heifers required treatment for which no health alert was generated by the CowManager system.
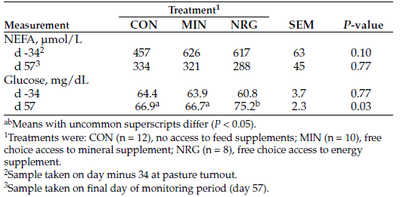
We found no differences (P > 0.10; Table 2) in initial or final serum NEFA concentrations among treatment groups. Initial serum glucose concentrations were not different (P = 0.77) among treatments. However, glucose concentrations were greater (P = 0.03) in NRG heifers, compared with CON and MIN heifers, at the end of the monitoring period.
We observed no differences in initial liver mineral concentrations among treatments (P > 0.13; Table 3). Liver mineral concentrations for Se were greater in NRG heifers, compared to CON and MIN heifers (P = 0.008). Iron concentrations were greater in NRG heifers, compared with CON and MIN heifers (P = 0.04). The NRG heifers had the highest concentrations of Co, then MIN heifers followed by lower concentrations in CON heifers (P < 0.001); however, copper (Cu), zinc (Zn), molybdenum (Mo) and manganese (Mn) were not different among treatments (P > 0.21).
According to guidelines published by Kincaid (2000), liver mineral concentrations for Fe, Zn, Mo and Mn are considered adequate for heifers among all treatment groups. Suggested adequate liver Cu concentrations have been defined as 125 to 600 micrograms per gram (μg/g) dry matter (DM) (Kincaid, 2000) or greater than 100 μg/g DM (Radostits et al., 2007). Therefore, heifers would be considered marginal (33 to 125 μg/g DM; Kincaid, 2000) to adequate or normal for liver Cu concentrations.
Selenium concentrations in the liver for heifers are considered adequate (1.25 to 2.50 μg/g DM; Kincaid, 2000). Liver Co levels at 0.08 to 0.12 μg/g DM or more indicate satisfactory Co status (McNaught, 1948), and heifers were above satisfactory levels. Overall, heifers in their respective treatment groups had adequate liver mineral concentrations.
In conclusion, the MCCC units were deployed successfully and serve as portable units that use solar power to run individual components and upload data to cloud-based data acquisition platforms. SmartFeed units were able to control intake of individual animals assigned to different treatments in a group pasture scenario.
Our results show that the feed-controlling portion of the MCCC can be used for precision feeding of individuals in expansive group-managed scenarios. The potential exists to develop targeted management strategies for cattle with distinct nutrient needs (high and low body condition scores or mixed groups of cows and heifers) while being managed in common pastures. The CowManager system was able to detect divergence in highly active behavior among treatment groups but also reported many false health and estrus-related alerts.
Table 1. Activity of heifers monitored using CowManager ear tags.
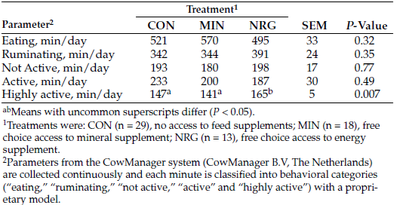
Table 2. Effects of mineral and mineral with energy supplements on serum metabolite concentrations in heifers grazing native range.
Acknowledgments
The authors acknowledge the staff at the Central Grasslands Research Extension Center for assistance with animal handling and forage collection. The authors also thank the North Dakota Agricultural Experiment Station (NDAES) Precision Agriculture Fund and the North Dakota State Board of Agricultural Research and Education Graduate Research Assistantship programs for their support for this effort.
Literature Cited
Bowman, J.G.P., and B.F. Sowell. 1997. Delivery method and supplement consumption by grazing ruminants: a review. J. Anim. Sci.75:543–550.
Cappellozza, B.I., R.F. Cooke, M.M. Reis, P. Moriel, D.H. Keisler and D.W. Bohnert. 2014. Supplementation based on protein or energy ingredients to beef cattle consuming low-quality cool-season forages: II. Performance, reproductive, and metabolic responses of replacement heifers. J. Anim. Sci. 92:2725–2734.
Cline, H.J., B.W. Neville, G.P. Lardy and J.S. Caton. 2009. Influence of advancing season on dietary composition, intake, site of digestion, and microbial efficiency in beef steers grazing a native range in western North Dakota. J. Anim. Sci. 87:375–383.
Cockwill, C.L., T.A. McAllister, M.E. Olson, D.N. Milligan, B.J. Ralston, C. Huisma and R.K. Hand. 2000. Individual intake of mineral and molasses supplements by cows, heifers and calves. Can. J. Anim. Sci. 80:681–690.
Kincaid, R.L. 2000. Assessment of trace mineral status of ruminants: A review. J. Anim. Sci. 77(Suppl E):1–10.
McNaught, K.J. 1948. Cobalt, copper and iron in the liver in relation to cobalt deficiency ailment. New Zeal J. Sci. Tech., Section A 30:26–43.
Patterson, J.D., W.R. Burris, J.A. Boling and J.C. Matthews. 2013. Individual intake of free-choice mineral mix by grazing beef cows may be less than typical formulation assumptions and form of selenium in mineral mix affects blood Se concentrations of cows and their suckling calves. Biol. Trace Elem. Res. 155:38–48.
Radostits, O.M., C.C. Gay, K.W. Hinchcliff and P.D. Constable. 2007. Veterinary Medicine: A textbook of the diseases of cattle, horses, sheep, pigs, and goats. Saunders Elsevier.
Schauer, C.S., K.K. Sedivec, M.L. Bauer, W.D. Slanger and G.P. Lardy. 2004. Self-limiting supplements fed to cattle grazing native mixed-grass prairie in the northern Great Plains. J. Anim. Sci. 82:298–306.
Tait, R.M., and L.J. Fisher. 1996. Variability in individual animal’s intake of minerals offered free-choice to grazing ruminants. Anim. Feed Sci. Technol. 62:69–76.
Performance of beef cattle overwintered on bale-grazed pasture or in a dry lot in south-central North Dakota
Michael Undi1 and Stephanie Becker1
1Central Grasslands Research Extension Center, NDSU
Allowing beef cattle to harvest their own forage potentially can decrease production costs by reducing reliance on inputs such as labor and machinery required for forage harvest. This study assesses the performance of beef cattle kept on pasture to bale graze or in dry-lot pens during the winter in North Dakota. Results show that bale grazing may be a viable alternative to keeping cattle in dry lots in the winter. Further, environmental conditions such as blizzards will not necessarily hinder bale grazing when proper precautions are taken to ensure that animals have access to water, feed and shelter.
Summary
The performance of beef cows managed in two overwintering environments (pasture or dry-lot pens) was assessed in a study conducted during three winters, from 2016 to 2018, at the Central Grasslands Research Extension Center, Streeter, N.D. Starting in the fall of each year, nonlactating pregnant Angus cows were divided into four groups of similar body weight (BW) and kept on pasture to bale graze or in dry-lot pens in the winter. Keeping cows on pasture or in dry-lot pens in the winter did not influence (P > 0.05) their final BW and final body condition score (BCS). However, daily gains were greater (P < 0.05) in bale-grazing cows relative to cows kept in a dry lot. Although both groups lost BCS, the loss was greater (P < 0.05) in cows kept in a dry lot. Results show that bale grazing is a viable alternative to keeping cattle in dry lots in the winter. Further, environmental conditions such as blizzards will not necessarily hinder bale grazing when proper precautions are taken to ensure that animals have access to water, feed and shelter.
Introduction
Winters in the northern Plains are characterized by cold temperatures, low wind chills, freezing rain and snow. A large portion of winter (40 to 70 days) averages minus 18 C, although extreme minimum temperatures of minus 51 C have been recorded (Enz, 2003).
The majority of beef cows in the northern Plains are housed in open dry-lot pens during the winter (Asem-Hiablie et al., 2016) and are exposed to these extreme winter conditions. In typical dry lots, cattle are fed mechanically harvested feeds such as hay and silage.
Winter feed costs, resulting from labor, machinery and energy required to provide feed, water and bedding to cattle kept in dry lots, make up more than 60% of total feed costs for most beef cow-calf operations (Taylor and Field, 1995). Thus, beef producers are interested in reducing winter feed costs by extending the grazing season.
Extending the grazing season by keeping cattle on pasture for a significant period of time during the winter allows animals to harvest their own food and decreases reliance on inputs such as machinery and energy required to harvest forage (D’Souza et al., 1990). By maximizing the use of grazed grass, the cheapest feed resource for ruminants (Hennessy and Kennedy, 2009), extending the grazing season can decrease production costs and enhance profitability of livestock production (D’Souza et al., 1990; Hennessy and Kennedy, 2009).
Strategies for extending the grazing season such as swath grazing, bale grazing and stockpiling have been evaluated (D’Souza et al., 1990; Willms et al., 1993; Volesky et al., 2002; McCartney et al., 2004; Jungnitsch et al., 2011; Kelln et al., 2011; Baron et al., 2014). The economic benefits from these strategies accrue mainly from cost reductions of feeds and feeding, labor, fuel, machinery maintenance and repair, and manure removal.
Environmentally, keeping cattle on pasture returns nutrients directly onto the land and allows for optimal nutrient capture by growing plants (Jungnitsch et al., 2011; Kelln et al., 2011). Depositing manure directly on pastures avoids nutrient accumulation in one place, minimizing nutrient loss to the environment through runoff or leaching (Kelln et al., 2012; Bernier et al., 2014).
Extending the grazing season must be assessed against benefits to the animal as well as to the producer. Local information on animal performance in extended grazing systems, especially bale grazing, as well as data on the economics of extended grazing under North Dakota winter conditions, is limited. Therefore, this study was conducted to assess the performance of pregnant beef cows managed in two overwintering environments (pasture or dry lot) under south-central North Dakota winter conditions.
Experimental Procedures
This study extended for three years, from 2016 to 2018. Starting in the fall of each year, nonlactating pregnant Angus cows (2016, n = 32, body weight [BW] = 599 ± 68 kilograms (kg); 2017, n = 40, BW = 620 ± 59 kg; 2018, n = 40, BW = 643 ± 47) were divided into four groups of similar BW and kept on pasture to bale graze or kept in dry-lot pens in the winter.
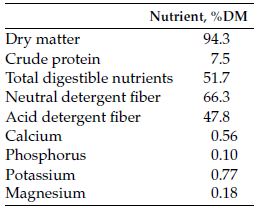
Pastured cows were kept in paddocks separated by three-strand, high-tensile wire electric fencing. Dry-lot pens contained a hay feeder and a water tank. Cows in both housing scenarios were offered the same Conservation Reserve Program (CRP) hay (Table 1), free choice.
Two-day body weights were taken at the start and end of the study. Two independent observers assigned body condition scores (BCS) using a 9-point system (1 = emaciated, 9 = obese; Wagner et al., 1988; Rasby et al., 2014) at the start and end of the study. Animal handling and care procedures were approved by the NDSU Animal Care and Use Committee.
Table 1. Nutrient composition of grass hay offered to cows bale grazing on pasture or kept in a dry lot.
Bale Grazing
Historically, the bale-grazing site was cropland in a corn and small-grain rotation. In the two years prior to the start of this study, the site was planted with cool-season cover crops, mainly rye and brassicas.
In 2016, the site was burned down with 2,4-D and Round-up in late April, after which meadow brome was planted in early May. Three-acre paddocks were separated using three-strand, high-tensile wire electric fencing. One water tank was placed between two paddocks. Wind breaks were placed in each paddock.
In early fall, round CRP hay bales (7.5% crude protein [CP]; 51.7% total digestible nutrients [TDN]) were placed in each paddock in two rows approximately 15 meters apart. Cows were allotted four bales in one grazing session; access to new bales was controlled using a single portable electric wire.
Cows were moved to a new set of four bales when the depth of waste feed remaining across the diameter of each bale was less than 10 centimeters (cm). Cows had ad libitum access to fresh water, mineral supplement and salt blocks.
Dry Lot
Two groups of cows were kept in dry-lot pens. Each pen contained a two-bale hay feeder and a Richie water tank. Dry-lot cows were fed the same CRP hay as the bale-grazed cows. Like the bale grazed cows, dry-lot cows had ad libitum access to fresh water, mineral supplement and salt blocks.
Results and Discussion
Animal Performance
Initial cow BW and BCS were similar (P > 0.05) between housing treatments in both years (Table 2). Keeping cows on pasture or in dry-lot pens in the winter did not influence (P > 0.05) final BW and final BCS. However, daily gains were greater (P < 0.05) in bale-grazing cows relative to cows kept in a dry lot.
Although both groups lost BCS, the loss was greater (P < 0.05) in cows kept in a dry lot (Table 2). Whether on pasture or in dry-lot pens, cows lost body weight and condition in the first year but maintained or gained weight and BCS in the second year (Figure 1).
Table 2. Performance of cows kept on pasture or in a dry lot in the winter.
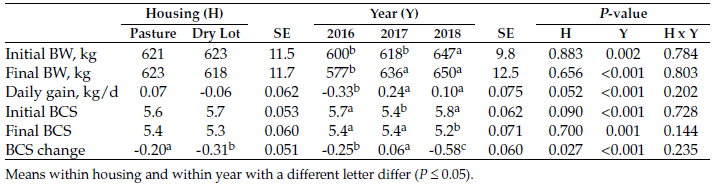
Figure 1. Average daily gains of cows (2016, n = 32; 2017, n = 40) kept on pasture or in a dry lot in the winter during two winters.
The first year of the study was marked by three blizzards, which led to huge snow accumulations. Despite snow depths being greater than 50 cm in selected places, cows were able to bale graze for 70 days before the termination of the study.
The study was terminated after accessing water points became impossible. This shows that strategies for extending the grazing season should be accompanied by a contingency plan for feed and water supplies in case grazing becomes impossible.
Here are some interesting observations from blizzard events of 2016: First, despite windbreaks, not all cows sought shelter during the blizzards. Some simply would stand on the leeward side of the bales, while other cows did not seek shelter at all and continued to graze.
Secondly, when water troughs were cleared of snow after each blizzard and refilled, not all cows visited the water troughs immediately, as anticipated. However, we observed what seemed to be a “catch up” period of several days following blizzards when water intake increased, as noted by more frequent filling of water troughs.
Events such as blizzards can prevent or drastically reduce access to water, requiring pastured cows to utilize snow as a source of water. Animals can survive on snow as shown in beef calves (Degen and Young, 1990a) and pregnant beef cows (Degen and Young, 1990b).
Cows in both housing scenarios lost body weight and condition in the first year, which was likely a combination of hay quality and environmental conditions. The hay was low in energy, protein and phosphorus (P) content and did not supply these nutrients to meet the requirements of cows in mid-gestation (National Research Council, 1996), particularly during adverse weather conditions as encountered in 2016. The positive animal performance in the second and third years may be attributed to more favorable environmental conditions.
Overall, daily gains were numerically greater in cows kept on pasture. The smaller-size dry-lot pens would be expected to give dry-lot cows a competitive energy expenditure advantage over cows on pasture. Animals on pasture spend more energy walking in search of food and water or shelter and more time eating and foraging for food than housed animals (Osuji, 1974).
Extra muscular activities, over and above those observed indoors, might increase maintenance energy requirements of animals on range by 25% to 50% (Osuji, 1974). However, this might not apply in bale-grazing situations where animals do not travel long distances to feed.
Keeping cattle on pasture or in dry-lot pens in the winter must be assessed against benefits to the animal, as well as financial benefits to the producer. Extending the grazing season reduces feed costs significantly because animals harvest their own food (D’Souza et al., 1990). Several studies (D’Souza et al., 1990; Willms et al., 1993; McCartney et al., 2004; Jungnitsch et al., 2011; Kelln et al., 2011; Baron et al., 2014) have shown economic advantages of extending the grazing season associated with reducing costs of feeds and feeding, labor, fuel, machinery maintenance and repair, and manure removal.
Results show that bale grazing is a viable alternative to keeping cattle in dry lots in the winter. Further, environmental conditions such as blizzards will not necessarily hinder bale grazing when proper precautions are taken to ensure that animals have access to water, feed and shelter.
Acknowledgments
Technical assistance provided by Dwight Schmidt, the late Rodney Schmidt, Cody Wieland, Thomas Mittleider and Rick Bohn is gratefully acknowledged.
Literature Cited
Asem-Hiablie, S., Rotz, C.A, Stout, R., Stackhouse-Lawson, K. 2016. Management characteristics of beef cattle production in the Northern Plains and Midwest regions of the United States. Prof. Anim. Sci. 32: 736–749.
Baron, V.S., Doce, R.R., Basarab, J., and Dick, C. 2014. Swath-grazing triticale and corn compared to barley and a traditional winter feeding method in central Alberta. Can. J. Plant Sci. 94: 1125–1137.
Bernier, J.N., Undi, M., Ominski, K.H., Donohoe, G., Tenuta, M., Flaten, D., Plaizier, J. C., Wittenberg, K.M. 2014. Nitrogen and phosphorus utilization and excretion by beef cows fed a low quality forage diet supplemented with dried distillers’ grains with solubles under thermal neutral and prolonged cold conditions. Anim. Feed Sci. & Technol. 193: 9–20.
Degen, A.A., and Young, B.A. 1990a. Average daily gain and water intake in growing beef calves offered snow as a water source. Can. J. Anim. Sci. 70: 711–714.
Degen, A.A., and Young, B.A. 1990b. The performance of pregnant beef cows relying on snow as a water source. Can. J. Anim. Sci. 70: 507–515.
D’Souza, G.E., Marshall, E.W., Bryan, W.B., and Prigge, E.C. 1990. Economics of extended grazing systems. Am. J. Alternative Agric. 5 (3): 120–125.
Enz, J.W. 2003. North Dakota topographic, climatic, and agricultural overview. www.ndsu.edu/fileadmin/ndsco/documents/ndclimate.pdf
Hennessy, D., and Kennedy, E. 2009. Extending the grazing season. Livestock. 14: 27–31.
Jungnitsch, P., Schoenau, J.J., Lardner, H.A., and Jefferson, P.G. 2011. Winter feeding beef cattle on the western Canadian prairies: impacts on soil nitrogen and phosphorous cycling and forage growth. Agric. Ecosyst. Environ. 141: 143–152.
Kelln, B.M., Lardner, H.A., McKinnon, J.J., Campbell, J.R., Larson, K., and Damiran, D. 2011. Effect of winter feeding system on beef cow performance, reproductive efficiency, and system cost. Prof. Anim. Sci. 27: 410–421.
Kelln, B., Lardner, H., Schoenau, J., and King, T. 2012. Effects of beef cow winter feeding systems, pen manure and compost on soil nitrogen and phosphorous amounts and distribution, soil density, and crop biomass. Nutr. Cycl. Agroecosyst. 92: 183–194.
McCartney D., Basarab, J.A., Okine, E.K., Baron, V.S., and Depalme, A.J. 2004. Alternative fall and winter feeding systems for spring calving beef cows. Can. J. Anim. Sci. 84: 511–522.
National Research Council. 1996. Nutrient requirements of beef cattle. 7th Revised Edition. National Academy Press, Washington, D.C.
Osuji, P.O. 1974. The physiology of eating and the energy expenditure of the ruminant at pasture. J. Range Manage. 27 (6): 437–443.
Rasby. R.J., Stalker, A., and Funston, R.N. 2014 Body condition scoring beef cows: A tool for managing the nutrition program for beef herds. University of Nebraska–Lincoln Extension. http://extensionpublications.unl.edu/assets/pdf/ec281.pdf
Taylor, R.E., and Field, T.G. 1995. Achieving cow/calf profitability through low cost production. Range Beef Cow Symposium. University of Nebraska, Lincoln. http://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1198&context=rangebeefcowsymp
Volesky, J.D., Adams, D.C., and Clark, R.T. 2002. Windrow grazing and baled-hay feeding strategies for wintering calves. J. Range Manage. 55: 23–32.
Wagner, J.J., Lusby, K.S., Oltjen, J.W., Rakestraw, J., Wetteman, R.P., and Walters. L.E. 1988. Carcass composition in mature Hereford cows: Estimation and effect on daily metabolizable energy requirement during winter. J. Anim. Sci. 66:603–612.
Willms, W.D., Rode, L.M., and Freeze, B.S. 1993. Winter performance of Hereford cows on fescue prairie and in drylot as influenced by fall grazing. Can. J. Anim. Sci. 73:881–889.

(Photo by Rick Bohn, NDSU)
Supplementation of beef cattle bale grazing grass hay in winter: effects on animal performance and soil nutrients
Michael Undi1 and Stephanie Becker1
1Central Grasslands Research Extension Center, NDSU
Ensuring that animals have adequate nutrition is important when bale grazing late in the season. For cows receiving poor-quality feed, this can be achieved by using supplementation methods that minimize labor and energy costs. This study examines methods of supplementing cows while bale grazing poor-quality hay. Results suggest that supplementation with good-quality alfalfa hay or a liquid supplement will not meet requirements of pregnant beef cows in early to mid-gestation in severely cold winters. Under such conditions, high-energy supplements such as corn dried distillers grains with solubles (DDGS) will be required to meet the nutrient shortfall. Supplementation with good-quality alfalfa hay or grass hay treated with a liquid supplement may be an option during mild winters.
Summary
Methods of supplementing beef cows bale grazing poor-quality grass hay were investigated in a study conducted during three winters, from 2016 to 2018, at the Central Grasslands Research Extension Center, Streeter, N.D. Methods evaluated were a) grass hay supplemented with good-quality alfalfa hay, b) grass hay supplemented with corn DDGS and c) grass hay treated with a liquid supplement. Results show that the method of supplementation depends on environmental conditions during the winter. In severely cold winters, good-quality alfalfa hay or a liquid supplement are not adequate to meet the requirements of pregnant beef cows in early to mid-gestation. Under such conditions, supplements such as corn DDGS will be needed to meet animal requirements. Supplementation with good-quality alfalfa hay or grass hay treated with a liquid supplement may be an option during mild winters.
Introduction
Beef cattle in the northern Plains typically graze poor-quality forages in the winter (Marshall et al., 2013). Poor-quality forages are generally low in energy, protein and minerals, impairing rumen microbial function, which leads to poor forage intake and digestion (Köster et al., 1996). The utilization of poor-quality forages can be improved through supplementation, which is especially important at critical times such as summer plant dormancy or fall and winter months (Caton and Dhuyvetter, 1997).
Cost-effective supplement delivery methods help minimize feed costs by reducing supplement delivery frequency (Schauer et al., 2005; Canesin et al., 2014; Gross et al., 2016) or eliminating pasture visits (Klopfenstein and Owen, 1981). Supplementation techniques that minimize or eliminate pasture visits in extended grazing systems will further the goal of minimizing winter feed costs.
This study was conducted to investigate methods of supplementing cows bale grazing poor-quality hay in the winter. The study examined beef cow performance and cost effectiveness of bale grazing supplementation strategies.
Experimental Procedures
This study extended for two years, from 2016 to 2017. Starting in the fall of each year, nonlactating pregnant Angus cows (2016, n = 64, body weight [BW] = 595 ± 65 kilograms [kg]; 2017, n = 80, BW = 621 ± 59 kg; 2018, n = 80, BW = 643 ± 45 kg) were divided into eight groups of similar total body weight and kept on bale-grazing pasture in the winter. The cows were pregnancy-checked prior to the start of the study to eliminate open cows. Cows were treated with IVOMEC (Ivermectin) pour-on during sorting.
The bale grazing site was a 26-acre field that historically was cropland, using a corn and small-grain rotation. In the two years prior to the commencement of this study, the site was planted to cool-season cover crops, mainly annual rye grass and brassicas.
The site was sprayed with 2,4-D and glyphosate in late April 2016 and seeded to a meadow brome grass, which was planted in early May 2016. The field then was divided into eight three-acre paddocks using three-strand, high-tensile wire electric fencing. One water tank was installed between two paddocks. The site was mowed prior to bale placement to reduce the possibility of cows grazing standing forage.
Forty round hay bales were placed in each paddock in two rows in the fall. Net wrap was removed prior to feeding. Bales were placed on their sides to reduce waste and loss of liquid supplement. Cows were allotted four bales at a time, and access to new bales was controlled using one portable electric wire. Windbreaks were placed in each paddock for protection.
Cows were assigned to one of four bale grazing treatments as follows: a) poor-quality hay (control), b) poor-quality hay supplemented with alfalfa hay, c) poor-quality hay supplemented with corn DDGS and d) poor-quality hay treated with a liquid supplement (Table 1). Poor-quality hay was obtained from a Conservation Reserve Program (CRP) field of mixed cool-season grasses that had not been harvested for several years.
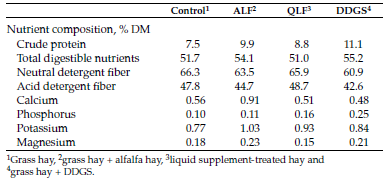
Table 1. Composition of grass hay and hay supplemented with alfalfa hay (ALF), a liquid supplement (QLF) or dried distillers grains with solubles (DDGS).
Cows supplemented with DDGS were fed 1.8 kg of DDGS/head/day twice weekly. Approximately 34 liters of liquid supplement (Quality Liquid Feeds Inc.) was poured onto upright bales. This amount of liquid supplement was calculated to increase hay protein content by approximately 3 percentage points.
Bales were allowed to sit upright after pouring until the supplement had seeped into the bale, after which the bales were flipped on their sides. One bale of alfalfa hay was fed for every three bales of poor-quality hay.
Cows had ad libitum access to water. Cows on the control, alfalfa hay and liquid supplement hay treatments were fed a 6-12+ mineral supplement (CHS Inc., Sioux Falls, S.D.). All cows were offered a salt block.
Two-day body weights were taken at the start and end of the study. Two observers assigned body condition scores (BCS) using a 9-point system (1 = emaciated, 9 = obese; Wagner et al., 1988; Rasby et al., 2014) at the start and end of the study. Animal handling and care procedures were approved by the NDSU Animal Care and Use Committee.
Soil samples were collected at two depths, 0 to 15 centimeters (cm) and 15 to 30 cm, and from three distance points: bale center, 10 feet from the bale center and 20 feet from the bale center. As well, soil samples were collected from bale grazed and ungrazed areas.
Results and Discussion
Initial cow BW and BCS were similar (P > 0.05) among treatments in both years. Final BW and BCS were not influenced (P > 0.05) by treatment. The diet by year interaction (P < 0.001) for daily gain (Table 2) shows that response to supplementation was dependent on the type of supplement used, as well as environmental conditions.
Table 2. Animal performance following bale grazing poor-quality grass hay supplemented with alfalfa hay, a liquid supplement or dried distillers grains with solubles (DDGS).
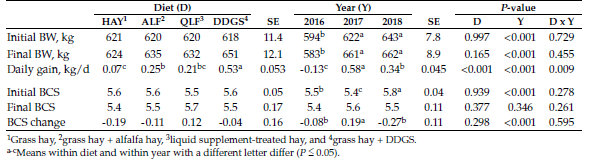
Figure 1. Cow daily grains following bale grazing grass hay supplement with alfalfa hay (alfalfa), a liquid supplement (QLF) or dried distillers grains with solubles (DDGS).
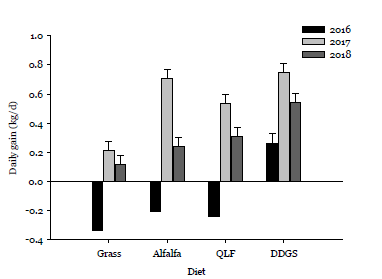
In the first year, cows supplemented with DDGS had positive daily gains, while supplementation with alfalfa hay or liquid resulted in weight loss (Figure 1). In the second year, more favorable environmental conditions resulted in similar performance in supplemented cows, whereas in the third year, DDGS supplementation clearly was superior to the other supplementation strategies (Figure 1).
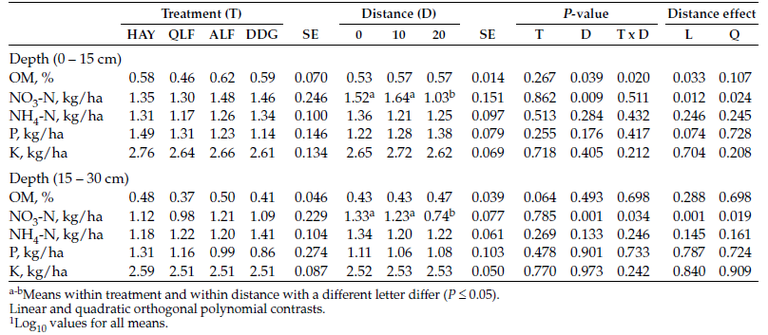
In the first year, supplementation did not influence (P > 0.05) soil organic matter, nitrate-N, ammonium-N, phosphorus (P) and potassium (K) at two soil depths (Table 3). As well, the distance from the center of the bale did not influence ammonium-N, P and K. However, nitrate-N content decreased linearly with increasing distance from the bale center. We found no difference in soil nutrients between bale grazed and ungrazed areas.
Table 3. Soil Concentration1 of organic matter, ammonium-N, nitrate-N, phosphours and potassium following bale grazing poor-quality hay supplements with DDGS, alfalfa hay or a liquid supplement.
During the first year of this study (2016), three blizzards occurred, which led to heavy snow accumulation in the paddocks. Despite snow depths greater than 20 inches in select places, cows were able to bale graze for 70 days before the termination of the study. The trial was terminated because cows no longer were able to reach the water source due to the heavy snowfall.
Environmental conditions will play a part in determining the success of supplementing cows bale grazing grass hay in the winter. When winters are harsh, poor-quality grass does not contain adequate amounts of energy, protein and phosphorus to meet nutritional requirement of cows in early to mid-gestation.
In the first year, cows supplemented with DDGS maintained BW and BCS, while cows supplemented with alfalfa hay or hay treated with a liquid supplement lost BW and BCS, suggesting that cow nutrient requirements were met by DDGS supplementation but not alfalfa hay or liquid supplementation. With more favorable winter conditions in the second year, supplementation with alfalfa hay, a liquid supplement or DDGS improved animal performance.
Results suggest that poor-quality grass hay does not contain adequate energy, protein and phosphorus to meet requirements of pregnant beef cows in early to mid-gestation when winters are severely cold. Under such conditions, supplementation with good-quality alfalfa hay or liquid supplement is not adequate and high-energy supplements such as corn DDGS will be required to meet the nutrient shortfall. Supplementation with good-quality alfalfa hay or grass hay treated with a liquid supplement may be an option during mild winters.
Acknowledgments
The excellent technical assistance provided by the late Rodney Schmidt, as well as Dwight Schmidt, Scott Alm, Megan Gross, Elisabeth Gnitka, Cody Wieland, Felipe Silva, Nico Negrin, Cheyanne Klein, Thomas Mittleider, Rick Bohn, Tom Lere (QLF) and Curt Lahr (QLF) is gratefully acknowledged.
Literature Cited
Asem-Hiablie, S., Rotz, C.A., Stout, R. Stackhouse-Lawson, K. 2016. Management characteristics of beef cattle production in the Northern Plains and Midwest regions of the United States. Prof. Anim. Sci. 32: 736–749.
Caton, J.S., and Dhuyvetter, D.V. 1997. Influence of energy supplementation of grazing ruminants: requirements and response. J. Anim. Sci. 75: 533–542.
Canesin, R.C., Berchielli, T.T., de Vega, A., Reis, R.A., Messana, J.D., Baldi, F., and Páscoa, A.G. 2014. Reducing supplementation frequency for Nellore beef steers grazing tropical forages. Sci. Agric. 71 (2): 105–113.
Gross, S.M., Neville, B.W., Brummer, F.A., and Undi, M. 2016. Frequency of feeding DDGS as a supplement to beef cows grazing corn residue. July 19-23. ASAS-ADSA-CSAS-WSASAS Joint Annual Meeting. Salt Lake City, Utah. Abstract # 612. Journal of Animal Science. Vol. 94, Suppl. http://academic.oup.com/jas/article/94/suppl_5/290/4766372
Kelln, B.M., Lardner, H.A., McKinnon, J.J., Campbell, J.R., Larson, K., and Damiran, D. 2011. Effect of winter feeding system on beef cow performance, reproductive efficiency, and system cost. Prof. Anim. Sci. 27: 410–421.
Klopfenstein, T., and Owen, F.G. 1981. Value and potential use of crop residues and by-products in dairy rations. J. Dairy Sci. 64: 1250–1268.
Köster, H.H., Cochran, R.C., Titgemeyer, E.C., Vanznat, E.S., Abdelgadir, I., and St. Jean, G. 1996. Effect of increasing degradable intake protein on intake and digestion of low quality, tall grass-prairie forage by beef cows. J. Anim. Sci. 74: 2473–2481.
Marshall, C.L., Fensterseifer, S.R., Arias, R.P., Funston, R.N., and Lake, S.L. 2013. The effect of winter protein supplementation during the third trimester on cow and subsequent calf performance. Proc. West. Soc. Am. Soc. Anim. Sci. 64: 103–105.
Rasby. R.J., Stalker, A., and Funston, R.N. 2014. Body condition scoring beef cows: A tool for managing the nutrition program for beef herds. University of Nebraska–Lincoln Extension. http://extensionpublications.unl.edu/assets/pdf/ec281.pdf
Schauer, C.S., Bohnert, D.W., Ganskopp, D.C., Richards, C.J., and Falck, S.J. 2005. Influence of protein supplementation frequency on cows consuming low-quality forage: performance, grazing behavior, and variation in supplement intake. J. Anim. Sci. 83: 1715–1725.
Wagner, J.J., Lusby, K.S., Oltjen, J.W., Rakestraw, J., Wetteman, R.P., and Walters. L.E. 1988. Carcass composition in mature Hereford cows: Estimation and effect on daily metabolizable energy requirement during winter. J. Anim. Sci. 66: 603–612.
Comparison of forage sampling strategies to monitor native range during the grazing season
Michael Undi1, Stephanie Becker1, Carl Dahlen2 and Kevin Sedivec1
1Central Grasslands Research Extension Center, NDSU
2Department of Animal Sciences, NDSU
Pasture forage sampling normally is accomplished by hand clipping. This two-year study compares hand clipping and rumen evacuation as forage sampling strategies to monitor changes in pasture chemical composition, forage intake and digestibility during the grazing season. Sampling strategy had the biggest impact on forage crude protein (CP) content and in vitro dry-matter digestibility (IVDMD), which were greater in samples collected through rumen evacuation. However, positive linear relationships between hand-clipped and masticate samples suggest that hand clipping provides reasonable estimates of pasture forage quality during the grazing season. Rumen evacuation may be a more suitable sampling strategy when a wide range of pasture conditions are anticipated.
Summary
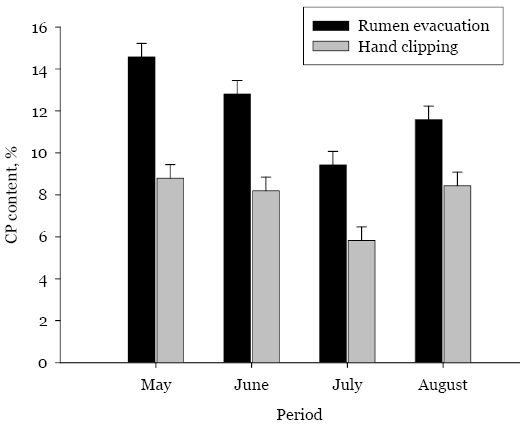
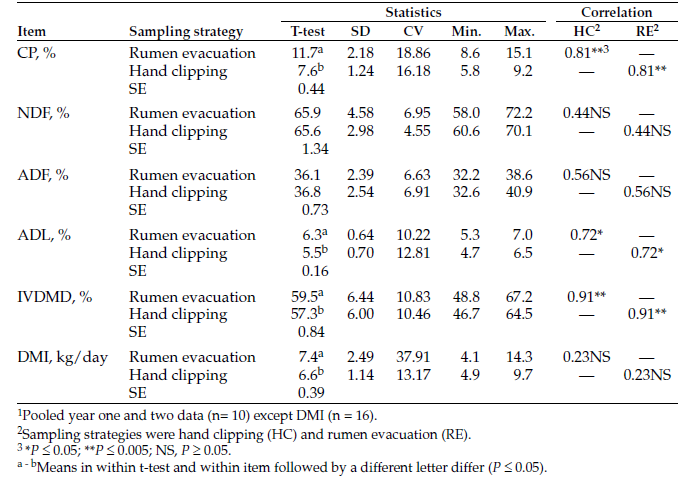
Hand clipping and rumen evacuation were compared as pasture forage sampling strategies to monitor changes in native range pasture chemical composition, forage intake and digestibility during the grazing season. Forage samples were collected by hand clipping or rumen evacuation in four periods in the first year and one period in the second year. Rumen evacuations were conducted with ruminal cannulated Angus heifers (n = 8, body weight [BW] = 597 ± 64 kilograms [kg], year one; n = 9, BW = 602 ± 76 kg, year two) that were kept in continuously-grazed pastures. Hand clipped and masticate samples were collected on the same day. Forage CP content was greater (P ≤ 0.05; 11.7 vs. 7.6 ± 0.44%) in masticate samples relative to clipped samples. Forage CP content declined (P ≤ 0.05) with advancing season. Regression analysis showed a significant relationship (r2 = 0.61; P ≤ 0.05) in CP content from the two sampling strategies. Forage neutral detergent fiber (NDF) and acid detergent fiber (ADF) content were not influenced (P > 0.05) by sampling strategy and increased linearly (P ≤ 0.05) with advancing season. Masticate samples had greater (P ≤ 0.05) acid detergent lignin (ADL) content relative to hand-clipped samples (6.3 vs. 5.5 ± 0.16%) and ADL contents from sampling strategies were correlated (r = 0.72; P ≤ 0.05). In vitro dry-matter digestibility (DMD) was greater (P ≤ 0.05) in masticate samples relative to hand-clipped samples and decreased linearly (P ≤ 0.05) with advancing season. Dry-matter intake estimated from masticate samples had a greater range (4.1 - 14.3 vs. 4.9 - 8.1 kg/day) and variability (CV; 37.7 vs. 13.2%) relative to estimates from hand clipped samples. Sampling strategy by period interaction (P ≤ 0.05) in dry-matter intake (DMI) showed that decline in DMI with advancing season was more pronounced in masticate samples relative to hand-clipped samples. Although sampling strategy had the biggest impact on forage CP content and IVDMD, hand clipping can provide useful estimates of pasture forage quality and nutrient changes during the grazing season, provided differences in forage chemical composition and IVDMD are considered. Sampling strategy should be taken into account when evaluating pasture quality.
Introduction
Beef producers in North Dakota depend largely on mixed-grass prairie as the primary forage source for cattle throughout much of the year (Johnson et al., 1998). Pasture productivity of mixed-grass prairie, measured by changes in crude protein content, fiber content and forage digestibility, declines with advancing season (Johnson et al., 1998; Cline et al., 2009; Cline et al., 2010), the rate of decline depending on time of the year, forage species and environmental conditions (McDowell, 1996).
Thus, pasture productivity should be monitored periodically so that decisions for optimal pasture use and cattle management are based on accurate information. In many cases, evaluation of pasture productivity is hindered by the inability to collect a representative sample mainly due to diverse plant communities and the rugged terrain of grazing lands (Holechek et al., 1982).
Pasture forage samples can be collected through several methods. Esophageal fistulation has been evaluated in several studies (Rama Rao et al., 1973; Vavra et al., 1978; Coffey et al., 1991; Olson, 1991) and provides the most representative sample of forage consumed by grazing animals (Holechek et al., 1982). Although sample collection through esophageal fistulation requires less labor, compared with other methods, esophageal fistulated cattle are expensive and difficult to maintain (Coffey et al., 1991).
Pasture sampling through rumen evacuation also has been used to obtain representative pasture samples (Olson, 1991; Hughes et al., 2010). Compared with esophageal fistulation, rumen fistulated animals are easier to maintain and representative samples can be collected during longer collection periods (Olson, 1991). Major limitations of rumen evacuation include time and labor to evacuate and clean the rumen, as well as depressed digestibility if evacuations are repeated frequently (Olson, 1991).
Nutritional management decisions for cattle often are based on pasture productivity estimated from hand-clipped forage samples (Hughes et al., 2010). Unlike sampling through esophageal fistulation or rumen evacuation, hand clipping requires less equipment and time and also produces sample free of salivary contamination (Holechek et al., 1982). However, hand clipping may misrepresent forage consumed because this strategy does not take into account diet selection (Holechek et al., 1982).
Despite these setbacks, clipping likely will remain the most practical strategy for pasture sampling. This study was conducted to compare hand clipping and rumen evacuation as sampling strategies for monitoring seasonal variation in forage intake, digestibility and forage chemical composition of mixed-grass prairie pastures.
Experimental Procedures
This two-year study was conducted at the North Dakota State University Central Grasslands Research Extension Center near Streeter, N.D. Animal handling and care procedures in this study were approved by the North Dakota State University Animal Care and Use Committee. The study was conducted with ruminal cannulated Angus heifers (n = 8, BW = 597 ± 64 kg, year one; n = 9, BW = 652 ± 91 kg, year two) that were kept in continuously grazed pastures and were co-grazed with cow-calf pairs (year one) and heifers (year two).
The grazing season was divided into four collection periods corresponding to May, June, July and August in the first year. Logistics resulted in late pasture turnout in the second year (2018), resulting in sample collection in July and August. Hand-clipped samples were collected by walking diagonally across pastures and hand clipping forage from 20 different locations in the pasture. The samples were clipped to a height of 3.75 centimeters (cm) above ground. Masticate samples were collected through a rumen evacuation technique described by Cline et al. (2010).
Results and Discussion
The effect of sampling strategy on pasture forage nutrients, intake and IVDMD in year one is shown in Table 1. Masticate samples had greater (P ≤ 0.05) CP content relative to hand-clipped samples. As well, forage CP content declined quadratically (P ≤ 0.05) with advancing season (Table 1). The tendency toward a sampling strategy by period interaction (P = 0.09) suggests that the magnitude of change in CP content with advancing season depended on sampling strategy (Figure 1).
Forage NDF and ADF content were not influenced (P > 0.05) by sampling strategy but increased linearly (P ≤ 0.05) with advancing season (Table 1). Hand-clipped samples had lower (P ≤ 0.05) ADL content relative to masticate samples (Table 1). Also, forage ADL content increased linearly (P ≤ 0.05) with advancing season.
Table 1. Pasture chemical composition, intake and digestibility estimated from forage samples collected by hand clipping or rumen evacuation (year).
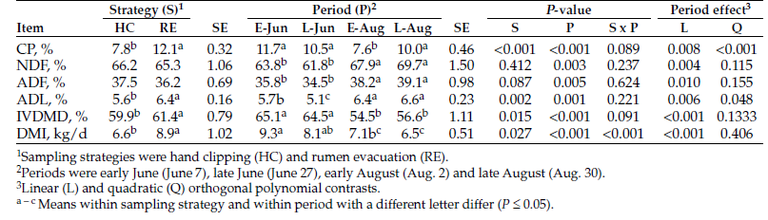
Figure 1. Change in forage CP content with advancing season estimated from forage samples collected by hand clipping or rumen evacuation.
In vitro DMD was greater (P ≤ 0.05) in masticate samples relative to hand-clipped samples. In vitro DMD decreased linearly (P ≤ 0.05) with advancing season (Table 1) but the rate of decline tended (P = 0.09) to depend on sampling strategy. The sampling strategy by period interaction (P ≤ 0.05) in DMI shows that the linear decline in DMI with advancing season was more pronounced in masticate samples relative to hand-clipped samples (Figure 2).
Late pasture turnout in the second year limited sampling to only one period (Table 2). Masticate samples had greater (P ≤ 0.05) CP content relative to hand-clipped samples. We found no differences (P > 0.05) in NDF, ADF and ADL content, as well as IVDMD, between masticate and hand-clipped samples.
Table 2. Pasture chemical composition and digestibility estimated from forage samples collected by hand clipping or rumen evacuation (year 2).
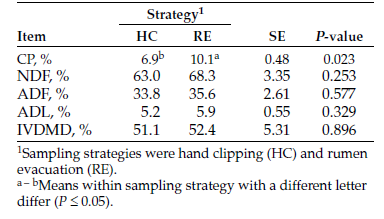
Figure 2. Change in DMI with advancing season estimated from forage samples collected by hand clipping or rumen evacuation.
When year one and two samples were pooled, forage CP content of masticate forage samples ranged from 8.6% to 15.1%, with a mean CP content of 11.7%, which was greater (P ≤ 0.05) than the CP content of hand-clipped samples (Table 3). Forage CP content resulting from the two sampling strategies were highly correlated (r = 0.81; P ≤ 0.05), and regression analysis showed a significant linear relationship (r2 = 0.61; P ≤ 0.05) between masticate and hand-clipped samples.
Pooled masticate and hand-clipped samples had similar (P > 0.05) forage NDF and ADF content (Table 3). Pooled masticate samples had greater (P ≤ 0.05) ADL content relative to hand-clipped samples (Table 3), but ADL content were correlated (r = 0.72; P ≤ 0.05; Table 3).
In vitro DMD of masticate and clipped samples were highly correlated (r = 0.91; P <0.005; Table 3), and regression analysis showed a significant linear relationship (r2 = 0.81; P ≤ 0.05) between the two sampling strategies. Dry-matter intake estimated from masticate samples had a greater range relative to estimates from hand-clipped samples (Table 3).
As well, DMI estimates from masticate samples were more variable as indicated by relatively greater CV. Dry-matter intakes estimated from the two sampling strategies were not correlated (Table 3).
Table 3. Pooled1 data comparing forage sampling strategies to estimate pasture chemical composition, dry-matter intake and digestibility.
This study compared rumen evacuation and hand clipping as sampling strategies to monitor seasonal nutritional quality changes of mixed-prairie pasture. Nutritional management decisions for cattle often are based on pasture productivity estimated from hand-clipped forage samples (Rama Rao et al., 1973; Dubbs et al., 2003; Hughes et al., 2010).
In this study, hand clipping followed normal practice as would be practiced by grazers, with no attempt to identify specific areas of the pastures where the animals were grazing. Hand clipping requires less equipment and time, and also produces samples free of salivary contamination (Holechek et al., 1982).
However, hand clipping may misrepresent forage consumed because this strategy does not take into account diet selection (Holechek et al., 1982). Despite these limitations, we anticipate that hand clipping will remain a method of choice for most grazers because it does not require cannulated animals.
Pasture sampling through rumen evacuation has been used to obtain representative pasture samples (Olson, 1991; Hughes et al., 2010) because samples collected represent forage consumed by animals. Rumen evacuation requires cannulated animals, and the process of collecting
masticate samples can be labor-intensive (Olson, 1991). In the present study, four individuals took at least six hours to collect masticate samples from nine cannulated heifers during each sampling cycle.
Forage CP content declined with advancing season as previously reported in the northern Great Plains (Johnson et al., 1998; Cline et al., 2009; Cline et al., 2010). The difference between sampling strategies was in magnitude of change in CP content (Figure 3). Forage CP content was 54% greater in masticate samples relative to clipped samples.
Studies that have reported greater CP content in esophageal-collected (Rama Rao et al., 1973; Coffey et al., 1991) or masticate samples (Dubbs et al., 2003; Hughes et al., 2010) relative to clipped samples attributed the lower CP content in clipped samples to failure of clipped samples to mimic grazing by not accounting for animal selection.
Coupled with a greater CP content range of masticate samples (8.6% to 15.1%), results from this study suggest that grazing cattle have the ability to select forage with higher CP content and will likely select forage with greater CP forages, even when forage quality is declining. This likely would occur only in cases where forage availability is not a limiting factor. A significant correlation between masticate and clipped CP values suggests that, despite limitations of clipping, this strategy still has value and can be used in situations where masticate samples cannot be obtained.
Contrary to studies that have reported greater NDF and ADF content in clipped samples relative to masticate (Dubbs et al., 2003; Hughes et al., 2010) samples, NDF and ADF contents of masticate and clipped samples in this study were similar. Differences between this and others studies could be due to clip sampling location where samples were collected in the same location as grazing animals or across pastures.
Forage NDF and ADF content increased with advancing grazing season, which previously has been reported in the northern Great Plains (Johnson et al., 1998; Cline et al., 2009; Cline et al., 2010) and reflects the association of advancing forage maturity with increased cell wall constituents (Van Soest, 1982).
Forage ADL content increased with advancing season mainly due to advancing forage maturity, which is associated with increased cell wall constituents, including lignin (Van Soest, 1982). The lower ADL content in clipped relative to masticate samples was unexpected and contrary to studies (Rama Rao et al., 1973; Coffey et al., 1991) that have shown greater ADL content in clipped samples.
In vitro DMD of masticate samples was greater than IVDMD from clipped samples and the two sampling strategies were highly correlated. A similar trend was reported in bahiagrass pastures where in vitro digestible organic matter (OM) of masticate samples was greater (60%) than hand-clipped samples (48.7%; Hughes et al., 2010). Decline in IVDMD with advancing grazing season also has been reported in other studies in the northern Great Plains (Johnson et al., 1998; Cline et al., 2009; Cline et al., 2010).
Typically, nutrition of ruminants grazing rangelands is complicated by diverse plant communities, changing topography and large seasonal and yearly variations in quantity and quality of available forage (Wofford et al., 1985). Precise estimation of forage intake by cattle on pasture depends on accurate determination of consumed forage components because chemical composition of consumed forage may differ from that of available forage resulting from animal selectivity and other processes involved with ingestion and mastication (Coffey et al., 1991). This is why determining a sampling strategy that provides a reliable estimate of consumed forage is important.
Forage samples for intake estimation can be obtained by following grazing animals for short durations and sampling grazed area (Wilson et al., 2010) or through rumen evacuation (Cline et al., 2010). A comparison of the two sampling strategies in this study shows that dry-matter intakes estimated from masticate samples were greater than estimates from hand clipped samples.
As well, masticate samples showed a greater DMI range and variability relative to clipped samples, indicating the ability of rumen evacuation to capture animal differences. Clearly, forage sampling strategy during individual animal DMI estimation on pasture will have an impact on intake estimations, and use of cannulated animals for forage sampling will provide more realistic DMI estimations.
This study suggests that hand clipping can provide reasonable estimates of pasture forage quality during the grazing season, provided you account for differences in CP, ADF content and IVDMD. Further, hand clipping can be a useful strategy in simple swards while rumen evacuation may be more suitable when a wide range of pasture conditions are anticipated.
Acknowledgments
The excellent technical assistance provided by the late Rodney Schmidt, as well as Dwight Schmidt, Scott Alm, Megan Gross, Elisabeth Gnitka, Cody Wieland, Felipe Silva, Nico Negrin, Cheyanne Klein and Rick Bohn is gratefully acknowledged.
Literature Cited
Coffey, K.P., Moyer, J.L., Lomas, L.W., and Turner, K.E. 1991. Technical note: sampling technique and drying method effects on chemical composition of tall fescue or fescue-ladino clover pasture samples. J. Anim. Sci. 69:423–428.
Cline, H.J., Neville, B.W., Lardy, G.P., and Caton. J.S. 2010. Influence of advancing season on dietary composition, intake, site of digestion, and microbial efficiency in beef steers grazing season-long or twice-over rotation native range pastures in western North Dakota. J. Anim. Sci. 88: 2812–2824.
Cline, H.J., Neville, B.W., Lardy, G.P., and Caton, J.S. 2009. Influence of advancing season on dietary composition, intake, site of digestion, and microbial efficiency in beef steers grazing a native range in western North Dakota. J. Anim. Sci. 87: 375–383.
Dubbs, T.M., Vanzant, E.S., Kitts, S.E., Bapst, R.F., Fieser, B.G., and Howlett, C.M. 2003. Characterization of season and sampling method effects on measurement of forage quality in fescue-based pastures. J. Anim. Sci. 81: 1308–1315.
Holecheck, J.L., Vavra, M., and Pieper, R.D. 1982. Methods for determining the nutritive quality of range ruminant diets: A Review. J. Anim. Sci. 54: 363–376.
Hughes, A.L., Hersom, M.J., Vendramini, J.M.B., Thrift, T.A., and Yelich, J.V. 2010. Comparison of forage sampling method to determine nutritive value of bahiagrass pastures. Prof. Anim. Sci. 26 (5): 504–510.
Johnson, H.M.A., Limb, R.F., Bauer, M.L., and Sedivec, K.K. 2018. Impacts of land management strategies on browse and nutritional quality of a resprouting shrub. Rangeland Ecology & Management (In Press).
Johnson, J.A., Caton, J.S., Poland, W., Kirby, D.R., and Dhuyvetter, D.V. 1998. Influence of season in dietary composition, intake, and digestion by beef steers grazing mixed-grass prairie in the northern Great Plains. J. Anim. Sci. 76:1682–1690.
McDowell, L.R. 1996. Feeding minerals to cattle on pasture. Anim. Feed Sci. Technol. 60: 247–271.
Olson, K.C. 1991. Diet sample collection by esophageal fistula and rumen evacuation techniques. J. Range Mgt. 44 (6): 515–519.
Rama Rao, M., Harbers, L.H., and Smith, E.F. 1973. Seasonal change in nutritive value of bluestem pastures. J. Range Mgt. 26(6): 419–422.
Van Soest, P.J. 1982. Nutritional ecology of the ruminant. O and B Books, Corvallis, Ore.
Vavra, M., Rice, R.W., and Hansen. R.M. 1978. A comparison of esophageal fistula and fecal material to determine steer diets. J. Range Mgt. 31 (1): 11–13.
Wilson, C., Undi, M., Tenuta, M., Wittenberg, K.M., Flaten, D., Krause, D.O., Entz, M.H., Holley, R., and Ominski, K.H. 2010. Pasture productivity, cattle productivity and metabolic status following fertilization of a grassland with liquid hog manure: A three-year study. Can. J. Anim. Sci.90: 233–243.
Wofford, H., Holechek, J. L., Galyean, M.L., Wallace, J.D., and Cardenas, M. 1985. Evaluation of fecal indices to predict cattle diet quality. J. Range Mgt. 38 (5): 450–454.
Effect of cover crop bale feeding (grazing) after native range or annual forage on steer grazing and feedlot performance, carcass measurements and carcass value
S. Şentürklü1,2, D.G. Landblom1 and S.I. Paisley3
1Dickinson Research Extension Center, NDSU
2Department of Animal Science, Çanakkale Onsekiz Mart University, BMYO, Çanakkale, Turkey
3Animal Science Department, University of Wyoming, Laramie, Wyo.
The objective of this grazing experiment, conducted within an integrated crop and livestock systems investigation, was to determine the value of feeding season-ending cover crop bales as a technique to extend the regular grazing season prior to feedlot entry to identify potential procedural effects on steer grazing performance and subsequently finishing performance, carcass measurements and carcass value. Steers grazing annual forage were heavier at the end of cover crop bale feeding, but not at the end of finishing. Feeding cover crop bales after grazing extended the period outside of the feedlot by 48.5 days. Treatment finishing performance did not differ resulting in identical finishing G:F, feed cost per pound of gain and total feedlot cost. Carcass quality grade was 97.9% Choice or better for both treatment groups and treatment gross carcass value was similar.
Summary
Ninety-six yearling steers of similar frame score (5.03) were assigned randomly to an extended grazing study to compare a control group of steers that grazed native range (NR) with steers that grazed a sequence of NR and annual forages (ANN: field pea-barley, corn, cover crop) to evaluate the effect of further extending the grazing season by feeding cover crop bales (bale grazing) on grazing performance and subsequently, finishing performance, carcass measurements, and economics. At the end of bale-feeding, the steers were finished in the feedlot. Extreme drought during the second year of the study (2017) affected the two-year averages causing large growth fluctuations for both NR and ANN forage steers. Native range steer gain declined as forage quality deteriorated, while ANN forage steers grazing unharvested corn gained significantly greater (P = 0.004). Compared to NR, ANN forage grazing plus cover crop bale feeding resulted in ANN steers gaining 0.40 pound per day more for the total 218.5-day grazing and bale feeding period, and ended 88 pounds heavier (P = 0.001). Live animal muscle and fat relationship was evaluated at the end of grazing. Steers that grazed NR had greater ribeye area per 100 pounds of carcass weight than ANN steers; however, ANN steers had greater percent intramuscular fat (IMF, P = 0.01). Marbling score was numerically greater for the ANN system steers, but did not differ (P = 0.10). Steers in the study were fed finishing diets for an average 104 days. Entering the feedlot 86 pounds heavier, the ANN steer growth was numerically slower than for the NR steers (P = 0.30); however, the ANN steer shrunk ending weight was 63 pounds heavier at the end of the finishing period. The NR steers tended to display compensating growth during finishing that resulted in identical ending gain:feed ratio and feed cost per pound of gain. Total feedlot cost per pound of gain did not differ. Grid carcass measurements identified ANN steers as having greater dressing percent (P = 0.01) and NR steers having greater muscling (REA:HCW, P = 0.02) and higher yielding carcasses (P = 0.01). Ending values for quality grade, grid market price per carcass hundredweight, and gross carcass value were similar and did not differ.
Introduction
A long-term (10-year) integrated crop and beef cattle investigation at the Dickinson Research Extension Center focuses on the interrelations of beef production, crop production, and soil health. For livestock integration, yearling steers provide the animal basis for vertical integration from birth to slaughter, replacing mechanical harvest with animal harvesting (Senturklu et al., 2017, 2018). Senturklu et al. (2018) compared economics of delayed feedlot entry to steers grown and finished in the feedlot and conducted a 10-Year (2003-2012) economic sensitivity analysis indicating that 70% of the time extended grazing and net return from delayed feedlot entry out performed traditional feedlot growing and finishing. Other extended grazing research has also shown improved forage quality, compensatory gain, lower breakeven cost and greater net profit (Lewis et al., 1990; Shain et al., 2005).
For the present study, a field pea-barley intercrop mix, silage-type corn, and a 13-species cover crop grown in the crop rotation preceded bale feeding of cover crop hay (12.8% crude protein) as a method for extending the grazing season after completion of NR and ANN forage grazing. The objective of this study was to determine the value of feeding season-ending cover crop bales as a technique for extending the period of time outside the feedlot on steer performance before feedlot entry and subsequently on finishing performance, carcass measurements and economics.
Experimental Procedures
The North Dakota State University Institutional Animal Care and Use Committee approved animal research procedures used in this study.
Ninety-six yearling crossbred steers (three reps of eight steers per pasture or field; frame score: 5.03) grazed western North Dakota native range (NR) or a forage sequence of NR and annual forages (ANN: field-pea barley mix, corn, and a 13-species cover crop mix). For bale grazing, a five-species cover crop hay was grown and baled in early July to obtain cover crop hay with a crude protein value of 12.8% (Table 1).
Treatment steers grazed NR as a common group from the first week of May until the third week of July, at which time the NR and ANN forage grazing treatments were separated. The NR steers continued to graze NR and the ANN treatment steers started grazing annual forages beginning with the field pea-barley mix, followed by unharvested corn, and ended with the 13-species cover crop. On Nov. 2 (year one) and Oct. 23 (year two), NR and ANN steer grazing ended and cover crop bale feeding started. Steers from NR and ANN forage treatments grazed cover crop bales in three 4.5 acre replicated fields per treatment.
Forage samples were collected using a 0.25-square-meter frame at the start and end of each forage grazing period. Forage samples were analyzed for CP, neutral detergent fiber (NDF), acid detergent fiber (ADF), in vitro dry matter disappearance (IVDMD), in vitro organic matter disappearance (IVOMD), calcium (Ca), phosphorus (P)and total digestible nutrients was calculated according to the following formula: (TDN = 81.38 + (CP % * 0.36) - (ADF % * 0.77).
Multiple blizzards, deep snow and drifting made feeding cover crop bales difficult the first year of the study and the extreme drought of 2017 reduced the number of cover crop grazing days. Therefore, cover crop hay was fed 41 days (year one) and 56 days (year two) before the steers were shipped for finishing to the University of Wyoming, SAREC feedlot, Lingle, Wyo.
Monitoring of live animal muscle and fat change during grazing, when steers were switched from one forage type to another, was conducted using an Aloka SSD-500V ultrasound machine and 3.5 Linear Array Transducer.
Steers were slaughtered at the Cargill Meat Solutions Plant, Ft. Morgan, Colo. Grid carcass measurements included hot carcass weight (HCW), fat depth (FD), ribeye area (REA), marbling score (MS), USDA yield grade (YG), quality grade (QG) and muscle-to-carcass weight ratio (REA: HCW) as well as calculated gross carcass value (HCW * Carcass Price/CWT).
Data were analyzed using the MIXED procedure of SAS (SAS Inst. Inc., Cary, N.C.). Means with P ≤ 0.05 differ significantly.
Results and Discussion
Nutrient analysis and calculated TDN values for the grazed forages and cover crop bales is shown in Table 1.
Steer grazing performance for NR and ANN forage crop sequence, and cover crop bale feeding is shown in Table 2. Steer growth for NR and ANN steers fluctuated during the grazing season due to drought, delayed cover crop seed germination that negatively impacted cover crop yield, and bale grazing blizzard conditions and deep snow THAT reduced the number of days bales could be fed.
Table 1. Nutrient analysis of grazed forges and cover crop bales.
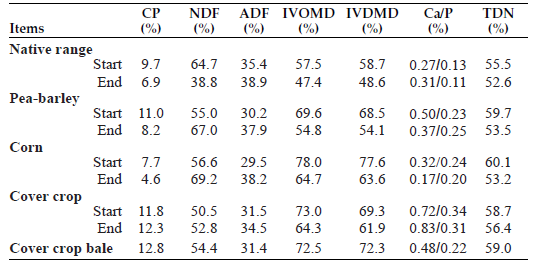
Table 2. Two-year effect of grazing system on yearling steer grazing performance.
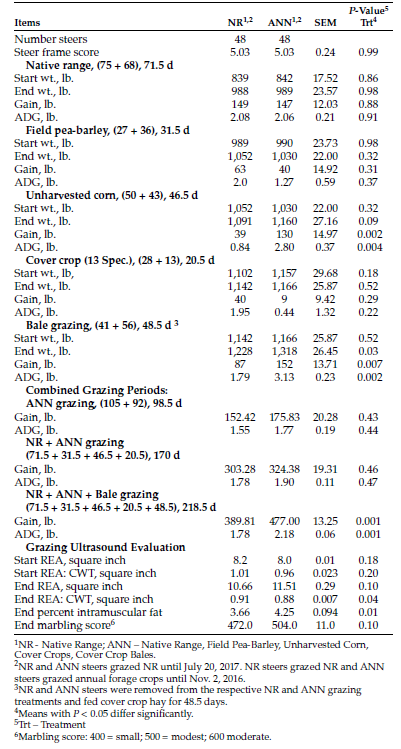
Grazing the sequence of NR and ANN forage sequence in the ANN treatment maintained ADG at approximately 1.90 pounds/day throughout the average 170-day grazing season compared; a 6.3% increase compared to the NR treatment (Table 2), which can be attributed to annual forage nutrient quality being consistently greater than NR forage (Table 1). The northern Great Plains growing season ends with the first killing frost in September (average date: Sept. 25); however, cover crop grazing continued until Nov. 2 (year one) and Oct. 23 (year two), when the NR and ANN forage grazing season ended.
At the end of the cover crop bale feeding period, ANN steers had greater ADG (P < 0.002) and weighed 90 pounds more (P = 0.03; Table 2) than the NR control steers. Although steer gain declined as NR matured with advancing season, fall rain stimulated range regrowth and steer gains recovered during September and October.
Cover crop bale feeding gain was 42.8% greater for ANN steers, compared with NR steers (P = 0.002). Given the somewhat slower growth commonly associated with extended grazing, a compensating gain response, such as the responses reported by Senturklu et al. (2017, 2018) and Choat et al. (2003), was expected. However, for the entire 218.5-day grazing season that included the bale-feeding period prior to feedlot entry, ANN steer gain compared to the NR control treatment was 18.3% better (Table 2).
Economically important muscle and fat tissue (REA and IMF), and marbling score were measured at the end of grazing with ultrasound (Table 2). Ribeye muscle area for ANN steers was increased during the 218.5-day grazing period but was not different compared to the NR control treatment (P = 0.10). The REA:CWT was greater for NR steers (P = 0.04). The percent of IMF was greater for ANN steers (P = 0.01); however, MS was not influenced by grazing treatment (P = 0.10).
Feedlot performance between the NR and ANN grazing treatments was similar (Table 3). ANN steers entered the feedlot weighing 6.8% more than NR control steers (P = 0.04), a margin of 86.0 pounds. The ending weight margin between treatments was 3.9% greater for the ANN treatment, a difference of 62.3 pounds, which did not differ significantly (P = 0.19).
Table 3. Systems two-year feedlot finishing performance of steers placed into feedlot after bale grazing.
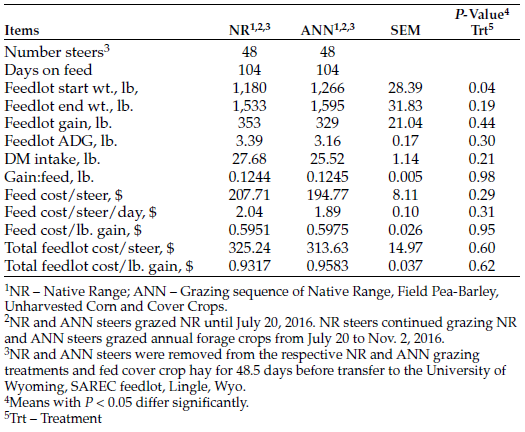
Differences between treatments for feedlot gain-to-feed (G:F) and feed cost/pound of gain were nearly identical (P = 0.96). Others have reported that delaying feedlot arrival by grazing NR and/or improved tame grass pastures resulted in greater feedlot starting body weight (BW) and ending slaughter weight, compared with steers that did not graze before entering the feedlot (Winterholler et al., 2008; Reuter and Beck, 2013; Lancaster et al., 2014), which was also documented by Senturklu et al. (2018).
Carcass dressing percent (P = 0.01) was greater for ANN steers. The NR control steers had greater muscling as indicated by the relationship between REA and HCW ratio for NR steers (P = 0.02), and NR steers also had more desirable USDA YG (P = 0.01) compared to ANN steers (Table 4). The ANN treatment steers consistently grazed higher-quality forage, and growth from cover crop hay increased the potential for fatter carcasses and more overweight carcass discounts. The muscling relationship identified for the NR steers at the end of grazing remained unchanged at the end of finishing.
Table 4. Two-year effect of grazing system on closeout carcass characteristics.
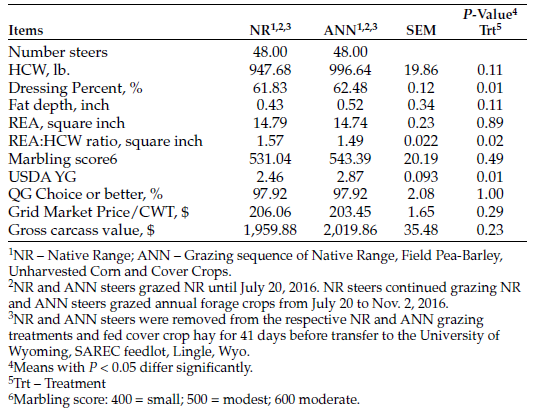
Gross carcass value was not influenced (P = 0.23) by treatment. Although carcass value margin was not great enough to identify a statistical difference in this study, results previously reported by Senturklu et al. (2017; 2018) show weight margins among groups entering the feedlot do not change appreciably by the end of the finishing period, and gross carcass value is routinely greater and more profitable for steers grazing ANN forages before feedlot entry, compared with NR.
Extending the period outside the feedlot feeding cover crop bales was an effective procedure for further delaying feedlot entry, which has been shown to increase final ending weight, HCW, quality grade, and gross carcass value. This study is being extended for an additional year to evaluate a larger data set.
Literature Cited
Choat, W.T., C.R. Krehbiel, G.C. Duff, R.E. Kirksey, L.M. Lauriault, J.D. Rivera, B.M. Capitan, D.A. Walker, G.D. Donart and C.L. Goad. 2003. Influence of grazing dormant native range or winter wheat pasture on subsequent finishing cattle performance, carcass characteristics, and ruminal metabolism. J. Anim. Sci. 81:3191-3201.
Lancaster, P.A., C.R. Krehbiel and G.W. Horn. 2014. A meta-analysis of effects of nutrition and management during the stocker and backgrounding phase on subsequent finishing performance and carcass characteristics. Prof. Anim. Sci. 30:602-612.
Lewis, J.M., T.J. Klopfenstein and R.A. Stock. 1990. Effects of rate of gain during winter on subsequent grazing and finishing performance. J. Anim. Sci. 68:2525-2529.
Shain, D.H., T.J. Klopfenstein, R.A. Stock, B.A. Vieselmeyer and G.E. Erickson. 2005. Evaluation of grazing alternative summer and fall forages in extensive beef cattle production systems. Prof. Anim. Sci. 21:390-402.
Reuter, R.R., and P.A. Beck. 2013. Southern Section Interdisciplinary Beef Cattle Symposium: Carryover effects of stocker cattle systems on feedlot performance and carcass characteristics. J. Anim. Sci. 91:508-515.
Senturklu, S., D.G. Landblom, R. Maddock, T. Petry and S. Paisley. 2017. Effect of retained ownership and vertical integration within an integrated cropping system among yearling steers of differing frame score on feedlot performance, carcass measurements, and systems economics following delayed feedlot entry. Proceedings, West. Sec., ASAS, Vol. 68:203-207.
Senturklu, S., D.G. Landblom, R. Maddock, T. Petry, C. Wachenheim and S. Paisley. 2018. Effect of yearling steer sequence grazing of perennial and annual forages in an integrated crop and livestock system on grazing performance, delayed feedlot entry, finishing performance, carcass measurements, and systems economics. J. Anim. Sci.
Winterholler, S.J., D.L. Lalman, M.D. Hudson, C.E. Ward, C.R. Krehbiel and G.W. Horn. 2008. Performance, carcass characteristics, and economic analysis of calf-fed and wheat pasture yearling systems in the southern Great Plains. Prof. Anim. Sci. 24:232-238.
The potential for dietary supplements commonly available in the upper Midwest to reduce frothy legume bloat based on in vitro degradation characteristics
Ronald J. Trotta1 and Kendall C. Swanson1
1Department of Animal Sciences, NDSU
The objective of this experiment was to compare the effects of different supplement sources on rate and extent of degradation and fermentation characteristics of an alfalfa-based diet using an in vitro fermentation system. Data from the current study indicate that soybean hull supplementation increases the extent of digestion while reducing the initial rate of degradation, suggesting that soybean hull supplementation has potential to reduce the incidence of frothy legume bloat. Supplements such as corn silage, grass hay and distillers grains may be effective at reducing bloat as well. Other supplements such as corn, beet pulp and distillers grains may increase diet digestibility at low levels of supplementation.
Summary
Two ruminally cannulated steers (2,046 ± 56.2 pounds body weight [BW]) were fed alfalfa hay at 2% of BW and served as donors of rumen fluid. Treatments were early bud alfalfa (CON) or a combination (dry-matter [DM] basis) of 80% early bud alfalfa with 20% of corn silage (CS), brome grass hay (BGH), soybean hulls (SBH), beet pulp (BP), corn grain (CORN), dried corn distillers grains with solubles (DDGS) or wheat middlings (MIDD). Tubes were incubated at multiple time points during a 72-hour period and assessed for rate and extent of digestion. Samples were run in duplicate and replicated for three days. The potential extent of DM degradation increased (P < 0.001) with CS, SBH, BP, CORN and DDGS supplementation, compared with CON. All supplements except MIDD decreased (P < 0.001) the fractional rates of DM degradation, compared with CON, with BGH, SBH and DDGS having the lowest rates. Supplementation of SBH had the lowest (P < 0.001) initial rate of DM degradation. These data indicate that SBH increases the extent of digestion while reducing the initial rate of degradation, suggesting that SBH has potential to reduce the incidence of frothy legume bloat.
Introduction
Alfalfa is the most widely grown perennial forage legume in the U.S. and can be used extensively in forage-based production systems for cattle and sheep. The primary factors limiting the use of high-quality alfalfa in cattle production systems are the 1) high and rapidly digestive protein fraction resulting in incomplete nitrogen (N) capture and 2) the incidence of frothy legume bloat.
Dietary supplementation is an effective mitigation strategy to reduce the incidence of legume bloat and improve animal performance (Wang et al., 2012). Many authors have reported that corn silage supplementation can reduce the incidence of frothy bloat when supplemented between 0.5 and 1 pound of DM per 100 pounds of BW per day (Bretschneider et al., 2007). These authors attributed this effect to reducing the initial rate of degradation of alfalfa. In addition, orchardgrass supplementation at 25% of dry-matter intake (DMI) has been shown to reduce legume bloat by more than 90% (Majak et al., 2008).
Although these supplements are effective at reducing frothy legume bloat, the mechanisms by which they reduce bloating are still unclear (Bretschneider et al., 2007). Moreover, forage-based supplements may not be feasible for all producers because of the infrastructure needed for harvest, storage and feeding.
Many supplement alternatives are available in the upper Midwest, and many byproducts may be more feasible and economically advantageous to producers. Therefore, the objective of this experiment was to compare the effects of different supplement sources on the rate and extent of degradation and fermentation characteristics of an alfalfa-based diet using an in vitro fermentation system.
Experimental Procedures
All animal procedures were approved by the North Dakota State University Institutional Animal Care and Use Committee.
An in vitro fermentation experiment was designed to evaluate the effects of supplementation on digestion parameters of alfalfa-based diets. Rumen fluid was collected from two ruminally cannulated steers fed alfalfa hay at 2% of BW. The rumen fluid was combined in a 1:4 ratio with buffer in 50 milliliter (mL) centrifuge tubes.
Freshly chopped vegetative to early bud alfalfa (CON) containing less than two buds per strand was used as the basal in vitro substrate. Seven supplement sources were tested in vitro: 1) corn silage (CS), 2) brome grass hay (BGH), 3) soybean hulls (SBH), 4) beet pulp (BP), 5) corn grain (CORN), 6) dried corn distillers grains with solubles (DDGS) and 7) wheat middlings (MIDD). Supplements were provided at 20% of the total DM.
A total of eight treatments were replicated two times daily and incubated for 0, 3, 6, 9, 12, 15, 18, 21, 24, 48 and 72 hours at 39 C. At each time point, each sample was run in duplicate and was replicated for three days. Samples were filtered through a Buchner funnel onto filter paper, dried overnight and weighed to determine the digestibility from the amount of DM remaining.
The potential rate and extent of in vitro DM and organic matter (OM) degradation was determined using the first-order asymptotic model (McDonald, 1981):
y = a + b (1 – e-c(t-L))
where y is the degradation after t hours, a is the rapidly degrading fraction, b is the slowly degrading fraction, c is the fractional rate of degradation, t is the incubation time (hours) and L is the lag time. Using the initial and final sample DM and OM masses, the absolute initial rate of degradation was calculated after three hours of incubation (Mertens, 2005). Data were analyzed using the PROC NLIN and PROC GLM procedures of SAS.
Results and Discussion
To evaluate the effects of different supplements for reducing frothy legume bloat, the rate and extent of digestion must be considered. The potential extent of DM degradation increased (P < 0.001) with CS, SBH, BP, CORN and DDGS supplementation, compared with CON. The greatest improvements in diet digestibility in this study were achieved with CORN or SBH supplementation. Soybean hulls and CORN increased (P < 0.001) the potential extent of DM degradation by 9% and 12.8%, respectively, compared with CON.
Table 1. Chemical composition (% of DM) of alfalfa hay (HAY), early bud alfalfa (CON), corn silage (CS), brome grass hay (BGH), soybean hulls (SBH), beet pulp (BP), corn grain (CORN), dried corn distillers grains with solubles (DDGS) or wheat middlings (MIDD) used for vitro fermentation substrates.
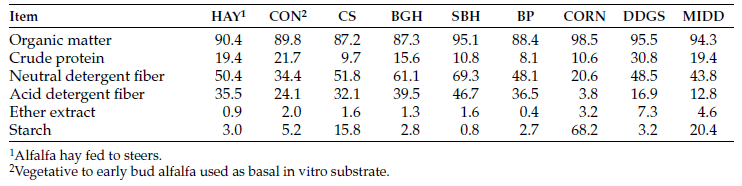
Table 2. Effect of supplemental corn silage (CS), brome grass hay (BGH), soybean hulls (SBH), beet pulp (BP), corn grain (CORN), dried corn distillers grains with solubles (DDGS) or wheat middlings (MIDD) on rate and extent of in vitro apparent dry matter degradation of early bud alfalfa.
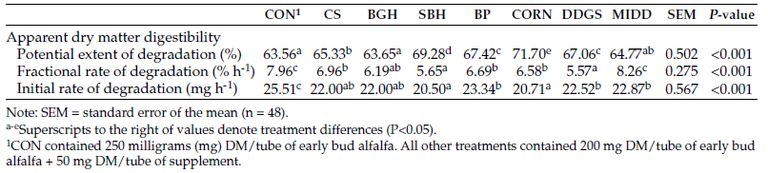
In general, researchers have demonstrated that at low levels of supplementation, CORN is a viable supplement in forage-based diets to improve performance in cattle (Matejovsky and Sanson, 1995; Trotta et al., 2018). However, at greater levels of supplementation, SBH may be a preferred supplement source because of less negative associative effects on forage intake and fiber digestibility.
Many factors may contribute to or exacerbate the incidence of frothy legume bloat, including the initial rate of degradation, grazing management, crop maturity, particle size, passage rate, ruminal pH and buffering capacity (Wang et al., 2012). Moreover, reductions in the occurrence of frothy legume bloat appear to be associated with a decrease in the initial rate of degradation (Wang et al., 2012). Howarth et al. (1982) suggested that a 25% to 30% reduction in the rate of degradation at six to eight hours was required to observe bloat-safe conditions.
In the current study, SBH and DDGS supplementation reduced the fractional rate of DM degradation by 29% and 30%, respectively, which may suggest that these supplements could have the potential to reduce frothy legume bloat in vivo. Brome grass hay supplementation was similar to SBH and DDGS in reducing the fractional rate of degradation, compared with CON. This supports previous reports by Majak et al. (2008), who demonstrated that orchardgrass hay supplementation can reduce the incidence of frothy legume bloat.
Likewise, the absolute initial rate of degradation measured after three hours of incubation was lower for SBH than DDGS, suggesting that SBH may be a more effective supplement at reducing bloat than DDGS. Because particle size was controlled in the current study, fibrous roughages such as BGH and CS may have lower rates of degradation in vivo. Therefore, future studies comparing the effects of BGH, CS and SBH on rates of in vivo ruminal degradation are warranted.
In conclusion, supplementation of SBH, DDGS or BGH reduced the fractional rate of DM and OM degradation greater than other supplements and CON. Likewise, SBH had the lowest absolute initial rate of degradation. The greatest increases in the potential extent of DM and OM degradation were observed with supplementation of SBH and CORN.
Based on in vitro degradation characteristics, this study provides evidence that SBH has the potential to reduce frothy legume bloat as an alternative to CS or BGH. More research is needed to determine if SBH is an effective supplement in vivo for reducing frothy legume bloat.
Acknowledgments
This work was partially supported by the Carl A. and Jean Y. White Memorial Endowment for Agricultural Research, a fund established by the NDSU Foundation and Alumni Association trustees.
Literature Cited
Bretschneider, G., M. Peralta, F.J. Santini, J.P. Fay and C. Faverin. 2007. Influence of corn silage supplementation before alfalfa grazing on ruminal environment in relation to the occurrence of frothy bloat in cattle. Anim. Feed Sci. Technol. 136:23-37.
Howarth, R.E., B.P. Goplen, S.A. Brandt and K.J. Cheng. 1982. Disruption of leaf tissues by rumen microorganisms: An approach to breeding bloat-safe forage legumes. Crop. Sci. 22:564-568.
Majak, W., G.J. Garland and T.J. Lysyk. 2008. The effect of feeding hay before fresh alfalfa on the occurrence of frothy bloat in cattle. Can. J. Anim. Sci. 88:29-31.
Matejovsky, K.M., and D.W. Sanson. 1995. Intake and digestion of low-, medium-, and high-quality grass hays by lambs receiving increasing levels of corn supplementation. J. Anim. Sci. 73:2156-2163.
McDonald, I. 1981. A revised model for the estimation of protein degradability in the rumen. J. Agric. Sci. 96:251-252.
Mertens, D.R. 2005. Rate and extent of digestion. Pages 805-851 in J. Dijkstra, J.M. Forbes and J. France, eds. Quantitative Aspects of Ruminant Digestion and Metabolism, 2nd ed. CAB International, Wallingford, UK.
Trotta, R.J., J.L. Klotz and D.L. Harmon. 2018. Effects of source and level of dietary energy supplementation on in vitro digestibility and methane production from tall fescue-based diets. Anim. Feed Sci. Technol. 242:41-47.
Wang, Y., W. Majak and T.A. McAllister. 2012. Frothy bloat in ruminants: Cause, occurrence, and mitigation strategies. Anim. Feed Sci. Technol. 172:103-114.
Maternal and Pre-weaning Nutrition
Maternal nutrient restriction of gestating ewes affects fetal digestive enzyme activities involved in carbohydrate and protein digestion
Ronald J. Trotta1, Manuel A. Vasquez-Hidalgo1, Kimberly A. Vonnahme1,2, and Kendall C. Swanson1
1Department of Animal Sciences, NDSU
2Zoetis Inc.
The objective of this experiment was to determine the effects of nutrient restriction of gestating ewes on fetal digestive enzyme activities in the pancreas and small intestine. Data from the current study demonstrate that maternal nutrient restriction decreases fetal pancreatic trypsin activity. Realimentation after maternal nutrient restriction during midgestation increases fetal lactase activity. Changes in fetal digestive enzyme activities potentially could influence post-natal performance.
Summary
To examine the effects of nutrient restriction during mid to late gestation on fetal digestive enzyme activities, 41 singleton ewes (48.3 ± 0.6 kilograms [kg] body weight [BW]) were assigned randomly to dietary treatments: 100% (control; CON; n = 20) or 60% of nutrient requirements (restricted; RES; n = 21) from day 50 until day 90 (midgestation). At day 90, 14 ewes (CON, n = 7; RES, n = 7) were euthanized. The remaining ewes were subjected to treatments of nutrient restriction or remained on a control diet from day 90 until day 130 (late gestation): CON-CON (n = 6), CON-RES (n = 7), RES-CON (n = 7) and RES-RES (n = 7) and were euthanized on day 130. The pancreas and small intestine were weighed, subsampled and assayed for digestive enzyme activity. Nutrient restriction during mid and late gestation did not affect fetal pancreatic or small intestinal mass. Maternal nutrient restriction during late gestation tended to decrease (p<0.08) fetal pancreatic trypsin activity. Sucrase activity was undetected in the fetal and maternal small intestine. Realimentation during late gestation after nutrient restriction during midgestation increased (p<0.05) lactase activity in the fetal small intestine. Fetal small intestinal maltase, isomaltase and glucoamylase did not respond to maternal nutrient restriction. Changes in maternal digestive enzyme activities are likely due to changes in pancreatic or small intestinal mass, as a function of feed intake.
Introduction
Maternal nutrition during gestation is a major determinant of fetal growth and development, and nutrient restriction during gestation can have adverse effects on maternal and fetal visceral tissues. Alterations in fetal visceral organ function during the pre-natal phase potentially can have negative effects on post-natal growth. Changes in maternal and fetal visceral organ mass in response to nutrient restriction during gestation potentially can alter maintenance energy requirements of the dam and fetus (Caton et al., 2019).
The pancreas and small intestine have important roles in post-ruminal nutrient digestion, and a limited amount of information is available on their function in response to nutritional adaptation. Specifically, both tissues produce carbohydrases, or glycohydrolases, that are digestive enzymes that hydrolyze glycosidic linkages of saccharides.
In nonpregnant ewes, nutrient restriction of ewes during mid to late gestation decreased maternal α-amylase activity but had no effect on fetal pancreatic digestive enzymes (Keomanivong et al., 2016). In beef cows, nutrient restriction during mid to late gestation increased fetal α-amylase activity but decreased trypsin activity (Keomanivong et al., 2017b).
No reports are available on the response of small intestinal carbohydrases (lactase, maltase, isomaltase, sucrase, glucoamylase) to nutrient restriction during pregnancy in ruminants. Therefore, the objective of this experiment was to determine the effects of nutrient restriction of gestating ewes on maternal and fetal digestive enzyme activities in the pancreas and small intestine.
Experimental Procedures
All animal care, surgical and sample collection procedures were approved by the North Dakota State University Animal Care and Use Committee.
Western whiteface ewes initially were placed on pasture with ad libitum access to hay and water, with two rams of proven fertility fitted with crayon marking harnesses. Mating was recorded every 12 hours. On day 30 of gestation, ewes were transferred to the North Dakota State University Animal Nutrition and Physiology Center. Singleton pregnant ewes were identified between day 30 and day 40 of gestation via Doppler ultrasonography.
The experimental design and description of treatment groups are presented in Figure 1. On day 50 of gestation, 41 ewes (BW = 48.3 ± 0.6 kg) were assigned to the following dietary treatments: 100% of nutrient requirements (control; CON; n = 20) or 60% of nutrient requirements (restricted; RES; n = 21) from day 50 to day 90 of gestation (midgestation).
At day 90, 14 ewes were euthanized (CON, n = 7; RES, n = 7) and the remaining ewes were subjected to treatments of nutrient restriction or remained on a control diet from day 90 until day 130 of gestation (late gestation; CON-CON, n = 6; CON-RES, n = 7; RES-CON, n = 7; and RES-RES, n = 7) and euthanized for sample collection.
On day 90 (n = 14) and day 130 (n = 27), ewes were euthanized with an overdose of sodium pentobarbital and the carcasses were eviscerated. Gastrointestinal tracts were removed and weighed, and digestive organs were separated for individual weights and subsample collection. The pancreas and small intestine were snap frozen in liquid nitrogen. Pancreatic samples were assayed for protein and α-amylase and trypsin activities. Small intestinal carbohydrases also were determined.
One unit (U) of enzyme activity equals 1 μmol substrate hydrolyzed per minute. Enzyme activity data are expressed as U/g tissue (concentration) and U/g protein (specific activity). Data were analyzed using the GLM procedure of SAS for effects of treatment and fetal sex.
Figure 1. Schematic of experimental design. Forty-one singleton ewes were fed a pelleted diet formulated to meet National Research Council (NRC) requirements (NRC, 1985) until day 50 of gestation. Dietary treatments were assigned from day 50 to day 90 (midgestation) and consisted of: 100% of nutrient requirements (control; CON; n = 20) or 60% of nutrient requirements (restricted; RES; n = 21) from day 50 to day 90 of gestation (midgestation). At day 90, 14 ewes were euthanized (CON, n = 7; RES, n = 7) and the remaining ewes continued on CON or RES until day 130, or CON ewes were RES from day 90 to 130, or RES ewes were realimented to CON from day 90 to 130. This resulted in four treatment groups: CON-CON (n = 6), CON-RES (n = 7), RES-RES (n = 7), RES-CON (n = 7). All remaining ewes on day 130 of gestation were euthanized for sample collection.
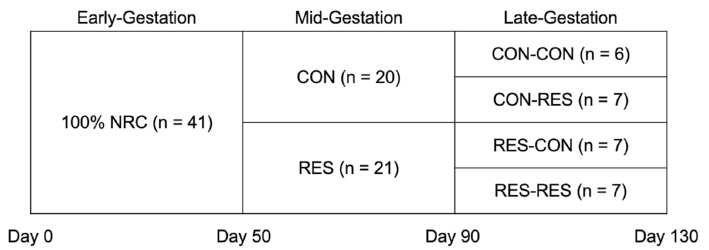
Results and Discussion
Fetal pancreas mass and small intestinal mass increased (P < 0.001) with day of gestation. Fetal small intestinal mass per kg of fetal BW increased from day 90 to day 130 (P = 0.04). Nutrient restriction during midgestation tended to decrease (P = 0.07) fetal small intestinal mass in tissues collected from day 90. In general, maternal pancreatic and small intestinal mass tends to change with plane of nutrition, while fetal pancreatic and small intestinal masses are less sensitive to changes in maternal diet (Keomanivong et al., 2017a).
In the fetal pancreas, pancreatic mass, total content of protein, α-amylase and trypsin increased (P < 0.003) with day of gestation. Interestingly, fetal trypsin activity decreased (P < 0.08) with maternal nutrient restriction during late gestation. Similarly, maternal nutrient restriction of beef cows during early or early to midgestation decreased trypsin-specific activity from fetal pancreas collected during late gestation (Keomanivong et al., 2017a).
Reasons for decreased trypsin activity in the fetal pancreas in response to nutrient restriction are unclear. What also is unclear is if trypsin activity in RES animals returns to CON levels post-partum.
Neville et al., (2010) reported that lambs from ewes restricted during mid and late gestation did not differ in postweaning total tract nitrogen (N) digestibility or N balance. Therefore, decreases in fetal trypsin activity may not affect the capacity for adequate protein digestion during post-natal life. Additional management or environmental factors during post-natal life potentially could mitigate negative effects of nutrient restriction during gestation.
Fetal brush border carbohydrases are imprinted early in development (Van Beers et al., 1995). Data from the current study demonstrate that fetal small intestinal carbohydrase activity is detectable (excluding sucrase) by the end of midgestation. What is unclear is how early in pre-natal development brush border enzyme activity is detectable in ruminants.
Table 1. Effects of nutrient restriction and stage of pregnancy of gestating ewes on fetal pancreatic mass, digestive enzyme activities and protein concentration.t
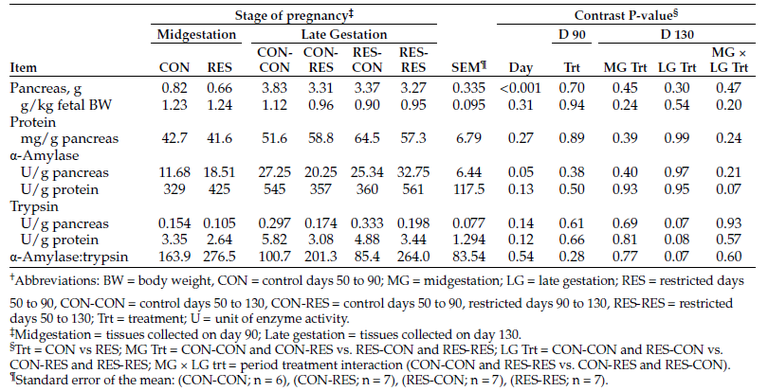
Table 2. Effects of nutrient restriction and stage of pregnancy of gestating ewes on fetal small intestinal mass, digestive enzyme activities and protein concentration.t
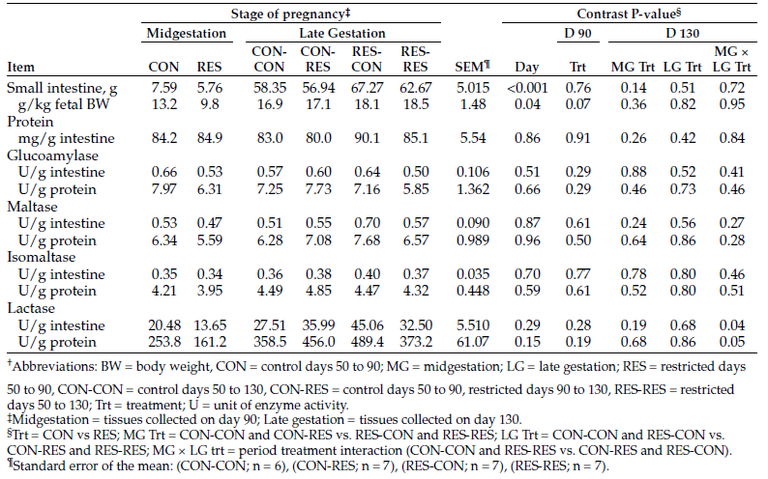
Fetal brush border α-glycohydrolases involved in small intestinal starch digestion were not affected by maternal nutrient restriction, suggesting that maternal nutrient restriction should not be used as a developmental programming strategy to increase fetal brush border α-glycohydrolase activities in sheep. Supporting these observations, Yunusova et al. (2013) found that offspring from nutrient-restricted ewes did not differ in jejunal maltase activity at day 180 post-partum.
Lactase produced in the mammalian intestine is the most important carbohydrase during early post-natal life because lactose is the primary carbohydrate ingested (Van Beers et al., 1995). In this study, we observed a midgestation × late-gestation interaction (P < 0.05) for lactase activity in the fetal small intestine.
Realimentation during late gestation after nutrient restriction during midgestation increased (P < 0.05) lactase activity in the fetal small intestine. This may indicate that nutrient restriction during midgestation potentially could be used as a programming strategy to increase fetal lactase activity.
However, the physiological consequences of increases in fetal lactase activity are not clear because greater than 90% of intestinal lactose supply is digested in the small intestine in neonatal ruminants. Regardless of treatment, fetal lactase activity was 60-fold greater on average than fetal maltase activity during late gestation.
In conclusion, maternal nutrient restriction during mid to late gestation causes alterations in fetal pancreatic and small intestinal digestive function. Fetal trypsin activity decreased with nutrient restriction during late gestation. Fetal brush border α-glycohydrolases involved in starch digestion were not influenced by nutrient restriction.
Realimentation during late gestation after nutrient restriction during midgestation increased lactase activity in the fetal small intestine. These data indicate that post-ruminal carbohydrases respond to nutrient restriction during mid and late gestation in ruminants.
Acknowledgments
This work was partially funded by U.S. Department of Agriculture-Agriculture and Food Research Initiative grant 2016-67016-24884.
Literature Cited
Caton, J.S., M.S. Crouse, L.P. Reynolds, T.L. Neville, C.R. Dahlen, A.K. Ward and K.C. Swanson. 2019. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 3:976-990.
Keomanivong, F.E., C.O. Lemley, L.E. Camacho, R.D. Yunusova, P.P. Borowicz, J.S. Caton, A.M. Meyer, K.A. Vonnahme and K.C. Swanson. 2016. Influence of nutrient restriction and melatonin supplementation of pregnant ewes on maternal and fetal pancreatic digestive enzymes and insulin-containing clusters. Animal. 10:440-448.
Keomanivong, F.E., L.E. Camacho, C.O. Lemley, E.A. Kuemper, R.D. Yunusova, P.P. Borowicz, J.D. Kirsch, K.A. Vonnahme, J.S. Caton and K.C. Swanson. 2017a. Effects of realimentation after nutrient restriction during mid- to late gestation on pancreatic digestive enzymes, serum insulin and glucose levels, and insulin-containing cell cluster morphology. J. Anim. Physiol. Anim. Nutr. 101:589-604.
Keomanivong, F.E., A.T. Grazul-Bilska, D.A. Redmer, C.S. Bass, S.L. Kaminski, P.P. Borowicz, J.D. Kirsch and K.C. Swanson. 2017b. The impact of diet and arginine supplementation on pancreatic mass, digestive enzyme activity, and insulin-containing cell cluster morphology during the estrous cycle in sheep. Dom. Anim. Endocrinol. 59:23-29.
Neville, T.L., J.S. Caton, C.J. Hammer, J.J. Reed, J.S. Luther, J.B. Taylor, D.A. Redmer, L.P. Reynolds and K.A. Vonnahme. 2010b. Ovine offspring growth and diet digestibility are influenced by maternal Se supplementation and nutritional intake level during pregnancy despite a common postnatal diet. J. Anim. Sci. 88:3645-3656.
Van Beers, E.H., H.A. Büller, R.J. Grand, A.W.C. Einerhand and J. Dekker. 1995. Intestinal brush border glycohydrolases: structure, function, and development. Crit. Rev. Biochem. Mol. Biol. 30:197-262.
Yunusova, R.D., T.L. Neville, K.A. Vonnahme, C.J. Hammer, J.J. Reed, J.B. Taylor, D.A. Redmer, L.P. Reynolds and J.S. Caton. 2013. Impacts of maternal selenium supply and nutritional plane on visceral tissues and intestinal biology in 180-day-old offspring in sheep. J. Anim. Sci. 91:2229-2242.
Effects of increasing amounts of supplemental leucine fed to neonatal Holstein bull calves on serum metabolites and amino acids, pancreatic hormones and intestinal enzymes
Jessica Reiners1, Michael Steele2, Kasey Maddock-Carlin1 and Kendall Swanson1
1Department of Animal Sciences, NDSU
2Department of Animal Biosciences, University of Guelph
The objective of this study was to determine the effects of increasing amounts of supplemental leucine on digestive enzymes in neonatal Holstein bull calves. Leucine supplementation decreased maltase and isomaltase activity in the intestine. Supplemental leucine during the neonatal phase can alter digestive enzyme activity, which may affect the digestibility of feeds as calves age.
Summary
Neonatal Holstein bull calves (n = 23) from a commercial dairy were used to determine the effects of increasing levels of supplemental leucine added to milk replacer fed to calves on pancreatic and intestinal digestive enzyme activity. Calves arrived in six blocks, with four calves per block. Calves were fed milk replacer at 2.5% of their body weight (dry-matter [DM] basis) with or without supplemental leucine for 28 days. Water was provided for ad libitum intake. On day 29, calves were euthanized, and samples of the pancreas and small intestine were collected. Supplementation of leucine did not affect pancreatic α-amylase (P > 0.27) or trypsin (P > 0.11) activities. Activity of glucoamylase in the small intestine was not affected (P > 0.41) by treatment. Isomaltase and maltase activity linearly decreased with increasing leucine supplementation (P < 0.05). Lactase (U/oz of intestine) responded quadratically (P = 0.02) with increasing leucine supplementation. Leucine supplementation to neonatal Holstein bull calves did not affect pancreatic enzyme activity. However, supplemental leucine did affect intestinal enzyme activity of calves. The long-term effects of leucine on intestinal enzyme activity are not known.
Introduction
In dairy cows and finishing cattle, starch is provided in diets as a major energy source for milk or meat production, but higher levels of inclusion can lead to ruminal acidosis and decreased performance. Starch that avoids degradation in the rumen is digested in the small intestine and could decrease the incidence of metabolic disorders.
In addition, starch digestion is 42% more efficient than in the small intestine (Huntington et al., 2006), and shifting starch degradation away from the rumen to the small intestine will capture greater energy without increasing risks of adverse metabolic effects. However, the small intestine is limited in its ability to degrade the available starch (Harmon, 2009), which can lead to feed waste. We hypothesize that nutrition during the neonatal period could influence the production of digestive enzymes later in life.
The pancreas is responsible for the secretion of α-amylase, which initiates starch digestion in the small intestine. Yu et al. (2014) showed that duodenal infusion of leucine (Leu) increased pancreatic α-amylase secretions in goats. Leucine has been shown to enhance the development of the small intestine of suckling piglets (Sun et al., 2015). Cao et al. (2019) reported that supplemental Leu fed to milk-fed calves had a limited effect on α-amylase secretions. Data on isomaltase and glucoamylase activity of neonatal calves are limited.
The objectives of this study are to determine the effects of different levels of supplemental leucine fed to neonatal calves on pancreatic and small intestinal digestive enzyme activity.
Experimental Procedures
All procedures involving the use of animals were approved by the North Dakota State University (NDSU) Institutional Animal Care and Use Committee.
Twenty-three neonatal Holstein bull calves (95.5 ± 2.56 pounds; 11.3 ± 0.57 days of age) were used. Calves were sourced from a commercial dairy and brought to the NDSU Animal Nutrition and Physiology Center. Calves were housed individually (2.99- by 3.94-foot pens) in a temperature-controlled room (72 F) on Tenderfoot flooring, and fed twice daily (7:30 a.m. and 4:30 p.m.) for 28 days.
Calves received milk replacer (Nurture, Cargill Inc., Minneapolis, Minn.) at 2.5% (DM basis) of their initial body weight (BW) split evenly between feedings. Milk replacer was reconstituted with 0.396 gallon of hot tap water at each feeding.
Calves were blocked by arrival date (n = four per block) into one of four treatment groups: milk replacer with no supplemental Leu (CON; n = 6), milk replacer with 0.04% Leu (LOW; n = 5; Ajinomoto, Raleigh, N.C.), milk replacer with 0.06% Leu (MED; n = 6), or milk replacer with 0.08% Leu (HIGH; n = 6). Daily Leu allotments were split evenly by feedings and mixed with milk replacer prior to feeding. Samples of the pancreas and a section of the jejunum in the small intestine were collected for hormone and enzyme analysis.
The experiment was a completely randomized block design. Calves were blocked by six arrival dates and assigned randomly to Leu treatment. All treatments were balanced based on age and weight of calves. Pancreatic and intestinal enzymes were analyzed using the GLM procedure in SAS (SAS 9.4, SAS Institute Inc., Cary, N.C.).
The experimental unit was calf, and the model included effects of block and treatment. Linear and quadratic effects of treatment were determined using orthogonal contrast statements. Significance was declared at P ≤ 0.05 and tendency at 0.05 < P < 0.10.
Results and Discussion
Activity of α-amylase was not affected by treatment (Table 1) when expressed as U/pancreas (P = 0.27) or U/lb BW (P = 0.34). Cao et al. (2018a,b) reported supplemental Leu can upregulate α-amylase secretion and can aid in post-ruminal starch digestion in mature ruminants. Cao et al. (2018a) reported a two-fold or greater increase in α-amylase activity vs. animals receiving control diets.
Our data does not support these results. These differences possibly are because of differences in daily delivery of Leu to animals. Our study offered Leu on a percent of BW of calves, whereas Cao et al. (2018a) offered Leu on a gram per day (g/d) basis. What is unclear is if Leu has a regulatory role in α-amylase activity of neonatal calves.
Trypsin was not affected by treatment when expressed as U/pancreas (P = 0.19) or U/lb BW (P = 0.11). Cao et al. (2018a) reported increases in pancreatic trypsin activity with dietary inclusion of Leu. However, Cao et al. (2018b) reported no differences in pancreatic activity of trypsin between control calves and Leu-supplemented calves. Our data showed that supplemental Leu had no effect on trypsin activity.
Glucoamylase activity was not affected (P > 0.41) by increasing Leu supplementation (Table 2). Data on neonatal glucoamylase activity are limited, and what is unclear is if Leu has a regulatory role on the activity of this enzyme.
Isomaltase activity linearly increased when expressed as U/oz intestine (P = 0.03). Reasons for these effects are not clear because data are limited on calf intestinal enzyme activity. However, decreasing activity from enzymes responsible for the breakdown of carbohydrates could cause a decrease in digestibility. What is unknown is if the activity of these enzymes remain suppressed when animals are transitioned to solid feed.
A quadratic effect of Leu on lactase activity on U/oz intestine was observed (P = 0.02) where Leu supplemented at LOW was less than all other treatments. Cao et al. (2019) reported no changes to lactase activity of milk-fed calves supplemented with Leu when compared with control calves. Leu may have limited to no regulatory role on lactase activity during the milk-feeding phase.
Increasing supplemental Leu linearly decreased maltase activity when expressed as U/oz intestine (P = 0.02). Radostits and Bell (1968) reported that the activity of maltase is very low in neonatal calves under 1 month of age. Our results showed an approximate 33% decrease in intestinal maltase activity of calves supplemented with Leu, when compared with control calves. However, because maltase activity is normally low in neonatal calves, the changes seen in our study may not continue once animals are transitioned to starch-based diets.
In conclusion, Leu supplementation during the neonatal period did not influence pancreatic digestive enzyme activity. However, supplemental Leu decreased intestinal enzyme activity. The long-term effects of these changes are not known.
Table 1. Pancreatic hormone analysis of calves fed milk replacer with or without supplemental leucine.
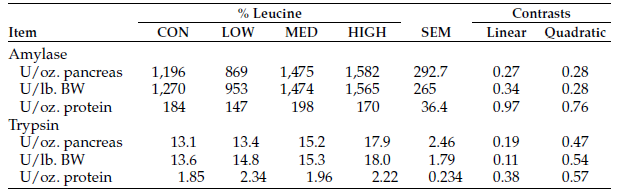
Table 2. Intestinal carbohydrate enzyme analysis of calves fed milk replacer with or without supplemental leucine.
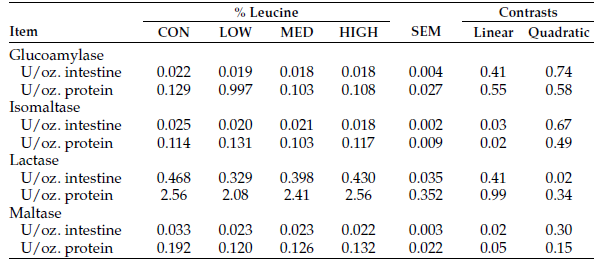
Acknowledgments
This project was supported by Animal Health and Production and Animal Products Accession No. 101206 from the U.S. Department of Agriculture’s National Institute of Food and Agriculture. The authors thank Cargill for supplying milk replacer and Ajinomoto for providing leucine used in this study. The authors also thank the employees of the NDSU Animal Nutrition and Physiology Center and the nutrition and physiology laboratories at NDSU.
Literature Cited
Cao, Y., S. Liu, X. Yang, L. Guo, C. Cai and J. Yao. 2019. Effects of leucine and phenylalanine on gastrointestinal enzyme activities in milk-fed Holstein dairy calves. Biosci. Reports. 39:BSR20181733.
Cao, Y., X. Yang, L. Guo, C. Zheng, D. Wang, C. Cai and J. Yao. 2018a. Regulation of pancreas development and enzymatic gene expression by duodenal infusion of leucine and phenylalanine in dairy goats. Livestock Sci. 216:9-15.
Cao, Y.C., X.J. Yang, L. Guo, C. Zheng, D.D. Wang, C.J. Cai, S. M. Liu and J.H. Yao. 2018b. Effects of dietary leucine and phenylalanine on pancreas development, enzyme activity, and relative gene expression in milk-fed Holstein dairy calves. J. Dairy Sci. 101:4235-4244.
Harmon, D.L. 2009. Understanding starch utilization in the small intestine of cattle. Asian-Aust. J. Anim. Sci. 22:915-922.
Huntington, G.B., D.L. Harmon and C.J. Richards. 2006. Sites, rates, and limits of starch digestion and glucose metabolism in growing cattle. J. Anim. Sci. 84:E14-24.
Radostits, O. M., and J.M. Bell. 1968. Nutrient digestibility by new-born calves fed milk replacer. Can. J. Anim. Sci. 48:293-302.
Sun, Y.Z. Wu, W. Li, C. Zhang, K. Sun, Y. Ji, B. Wang, N. Jiao, B. He, W. Wang, Z. Dai and G. Wu. 2015. Dietary L-leucine supplementation enhances intestinal development in suckling piglets. Amino Acids. 47:1517-1525.
Yu, Z.P., M. Xu, F. Wang, K. Liu, J.H. Yao, Z. Wu, D.K. Qin and F.F. Sun. 2014. Effect of duodenal infusion of leucine and phenylalanine on intestinal enzyme activities and starch digestion in goats. Livest. Sci. 162:134-140.
Effects of increasing amounts of supplemental leucine fed to neonatal Holstein bull calves on growth, digestibility, and muscle and organ weights during the milk-feeding phase
Jessica Reiners1, Michael Steele2, Kasey Maddock-Carlin1 and Kendall Swanson1
1Department of Animal Sciences, NDSU
2Department of Animal Biosciences, University of Guelph
The objective of this study was to determine the effects of graded amounts of supplemental leucine on growth and nutrient digestibility of neonatal bull calves. Leucine did not affect live calf performance during the 28-day period but affected pancreas weight. Short-term effects of leucine were limited, but long-term effects are unknown.
Summary
Neonatal Holstein bull calves (n = 23) from a commercial dairy were used to evaluate the effects of differing levels of supplemental leucine added to milk replacer fed to calves on growth, muscle weights and organ weights. Calves arrived in six blocks, with four calves per block. Calves were fed milk replacer at 2.5% of their body weight with or without supplemental leucine for 28 days. Water was provided for ad libitum intake. Body weights of calves were recorded on days zero, 1, 18, 22, 28 and 29. From days 22 to 29, calves were housed in metabolism stalls for total collection of urine and feces to determine digestibility of milk replacer. On day 29, calves were euthanized to determine the effects of leucine on organ growth and development. Calf weight and average daily gain (ADG) did not differ at any point during the experimental period (P > 0.15). Digestibility also was unaffected (P > 0.54) by treatment for dry matter (DM), organic matter (OM), crude protein (CP), ash or nitrogen (N). Further, nitrogen retained was not different and averaged 0.880 ounce per day (P = 0.59). Select organ weights were affected by treatment, including a linear increase in pancreas weight (P = 0.05), and quadratic effects on omasum (P = 0.01) and spleen (P = 0.02) weights. Leucine supplied to neonatal Holstein bull calves did not affect body weight or nutrient digestion of milk replacer but did influence pancreatic weight and weights of the omasum and spleen. Long-term effects of neonatal leucine supply to calves are unknown.
Introduction
Skeletal muscle protein synthesis is very rapid during the neonatal period, which relies heavily on amino acid (AA) intake. However, AA requirements of milk-fed calves are not well understood, but an imbalance in AA supply, or an oversupply of AA that is not limiting for growth, can decrease growth potential. Leucine (Leu) is known as one of the main AA involved in skeletal muscle protein synthesis, along with other roles in metabolism.
Leucine is known to be an essential AA because the body cannot supply adequate amounts to meet bodily needs. Leucine typically is not considered to be limiting in typical diets fed to ruminants, but it is a functional AA that aids in regulating key metabolic pathways for growth and maintenance.
Additionally, Leu stimulates insulin secretion and the mammalian target of rapamycin (mTOR), both regulators of muscle protein synthesis. Escobar et al. (2010) and Columbus et al. (2015) showed that Leu supplementation increased protein synthesis and body weight in neonatal piglets.
Data are limited on the effects of supplemental Leu fed to neonatal calves. The objective of this study was to determine the effects of short-term feeding of milk replacer with increasing amounts of supplemental Leu fed to neonatal calves on body weight, digestibility of milk replacer, nitrogen retention and organ weights.
Experimental Procedures
All procedures involving the use of animals were approved by the North Dakota State University (NDSU) Institutional Animal Care and Use Committee.
Twenty-three neonatal Holstein bull calves (95.5 ± 2.56 pounds; 11.3 ± 0.57 days of age) were used. Calves were sourced from a commercial dairy and brought to the NDSU Animal Nutrition and Physiology Center. Calves were housed individually (2.99- by 3.94-foot pens) in a temperature-controlled (72 F) room on Tenderfoot flooring, and fed twice daily (7:30 a.m. and 4:30 p.m.) for 28 days.
Calves received milk replacer (Nurture, Cargill Inc., Minneapolis, Minn.) at a 2.5% dry-matter basis of their initial body weight (BW) split evenly between feedings. Milk replacer (Table 1) was reconstituted with 0.396 gallon of hot tap water at each feeding.
Calves were blocked by arrival date (n = four per block) into one of four treatment groups: milk replacer with no supplemental Leu (CON; n = 6), milk replacer with 0.04% Leu (LOW; n = 5; Ajinomoto, Raleigh, N.C.), milk replacer with 0.06% Leu (MED; n = 6) or milk replacer with 0.08% Leu (HIGH; n = 6). Daily Leu allotments were split evenly by feedings and mixed with milk replacer prior to feeding. Samples of reconstituted milk replacer were taken for analyses.
Table 1. Nutrient profile of reconstituted calf milk replacer.
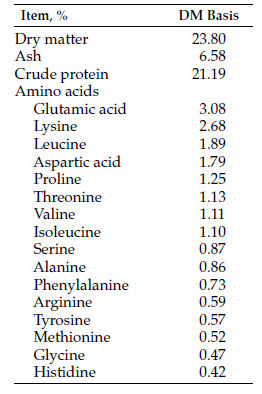
Calves were weighed for two days (day zero and day 1) at the beginning of the experiment, and on days 18, 22 and 28, and immediately prior to tissue collection on day 29. On day 29, calves were weighed and stunned via captive bolt before exsanguination for tissue collection.
The experiment was a completely randomized block design. Calves were blocked by six arrival dates and assigned randomly to Leu treatment. All treatments were balanced based on the age and weight of calves.
The nutrient profile of consumed milk replacer and organ weight was analyzed using the GLM procedure in SAS (SAS 9.4, SAS Institute Inc., Cary, N.C.). The experimental unit was calf, and the model included effects of block and treatment.
Linear and quadratic effects of treatment were determined using orthogonal contrast statements. Linear and quadratic effects of treatment were determined by orthogonal contrasts. Significance was declared at P ≤ 0.05 and tendency at 0.05 < P < 0.10.
Results and Discussion
Neither initial nor final calf weight were affected by treatment, and weights were not different among treatments throughout the study (Table 2). Average daily gain also was not influenced by treatment (data not shown).
Nutrient digestibility did not differ among treatments (Table 3). Nitrogen retention also did not differ among treatments.
Weights of omasum and spleen showed a quadratic effect (P = 0.01 and P = 0.02, respectively) of treatment (Table 4). Pancreas weights increased linearly with treatment (P = 0.05). No other weight measurements of organs or muscles were affected by Leu supplementation.
Table 2. Weight (pounds) of calves fed milk replacer with or without supplemental leucine.
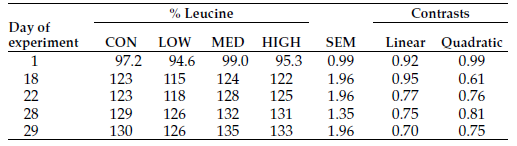
Table 3. Digestibility of nutrients and nitrogen retained in calves fed milk replacer with or without supplemental leucine.
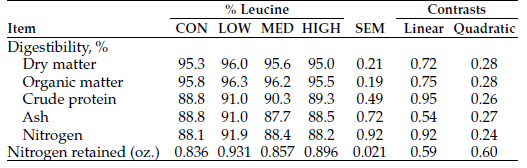
Table 4. Organ weights of calves fed milk replacer with or without supplemental leucine.
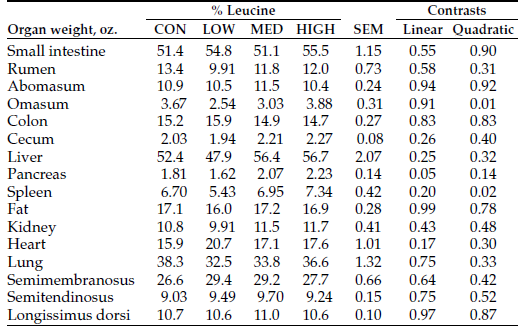
Columbus et al. (2015) reported that supplemental Leu to neonatal pigs increased piglet growth and muscle development. Cao et al. (2018) reported that dietary inclusions of supplemental Leu to Holstein calves did not affect final BW, similar to our results. Our results, coupled with that of Cao et al. (2018), suggest that Leu supplementation may not be effective in increasing muscle development in calves but may be effective in neonatal pigs.
The effects of leucine supplementation on nutrient digestibility in neonatal calves is unknown. Our results indicate that the digestibility and nitrogen retention of the milk replacer was high but was not influenced by Leu supplementation. The observed digestibility values were greater than that reported by Radostits and Bell (1968), and similar to that reported by Blaxter and Wood (1951).
Numerous advances have been made in milk replacer formulation. More recent data is limited on calf nutrient digestibility when fed diets containing milk replacer with no dry starter feed.
Organ weights of calves in this study were not influenced by Leu supplementation, with the exception of select organs. The differences in pancreatic weight could influence digestive enzymes and insulin production. These potential changes may affect the ability to break down and absorb nutrients from feed, and changes to insulin production could increase mTOR activity. In addition, the effects of Leu supplementation increased weights of the omasum and spleen; however, the physiological effects of these changes are not well defined.
In conclusion, Leu supplementation during the neonatal period had no effects on the growth and digestion of calves when fed for 28 days. Increasing Leu supplementation did affect pancreatic weight, but long-term effects of these weight differences are not known.
Acknowledgments
This project was supported by Animal Health and Production and Animal Products Accession No. 101206 from the U.S. Department of Agriculture’s National Institute of Food and Agriculture. The authors thank Cargill for supplying milk replacer and Ajinomoto for providing leucine used in this study. The authors also thank the employees of the NDSU Animal Nutrition and Physiology Center; the nutrition, physiology and muscle biology laboratories at NDSU; and students who assisted with this project.
Literature Cited
Blaxter, K., and W. Wood. 1951. The nutrition of the young Ayrshire calf. Br. J. Nutr. 5:11-25.
Cao, Y.C., X.J. Yang, L. Guo, C. Zheng, D.D. Wang, C.J. Cai, S.M. Liu and J.H. Yao. 2018. Effects of dietary leucine and phenylalanine on pancreas development, enzyme activity, and relative gene expression in milk-fed Holstein dairy calves. J. Dairy Sci. 101:4235-4244.
Columbus, D.A., J.S. Wagner, A. Suryawan, H.V. Nguyen, A.H. Garcia, M.L. Fiorotto and T.A. Davis. 2015. Impact of prolonged leucine supplementation on protein synthesis and lean growth in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 309:E601-610.
Escobar, J., J.W. Frank, A.Suryawan, H.V. Nguyen, C G. Van Horn, S.M. Hutson and T.A. Davis. 2010. Leucine and α-ketoisocaproic acid, but not norleucine, stimulate skeletal muscle protein synthesis in neonatal pigs. J. Nutr. 140:1418-1424.
Radostits, O.M., and J.M. Bell. 1968. Nutrient digestibility by new-born calves fed milk replacer. Can. J. Anim. Sci. 48:293-302.
Rogers, Q.R., and A.R. Egan. 1975. Amino acid imbalance in the liquid-fed lamb. Aust. J. Biol. Sci. April 28(2):169-181.
Schäff, C., J. Gruse, J. Maciej, M. Mielenz, E. Wirthgen, A. Hoeflich, M. Schmicke, R. Pfuhl, P. Jawor, T. Stefaniak and H. Hammon. 2016. Effects of feeding milk replacer ad libitum or in restricted amounts for the first five weeks of life on the growth, metabolic adaptation, and immune status of newborn calves. PLoS One. 11:1-24.
Williams, A.P., and D. Hewitt. 1979. The amino acid requirements of the preruminant calf. Br. J. Nutr. 41:311-319.
Effects of supplemental dietary leucine on lamb growth performance when provided during the milk-feeding phase
Jessica Reiners1, Travis Hoffman1 and Kendall Swanson1
1Department of Animal Sciences, NDSU
The objective of this study was to determine long-term effects of supplemental leucine fed to neonatal lambs on pre- and postweaning growth, organ weight and carcass characteristics. Lambs receiving leucine had a 10-pound increase in weaning weight, compared with controls, but no difference in finished weight or carcass characteristics. Supplemental leucine may increase producer profitability of weaned lambs with no detrimental effects on final products.
Summary
The objective of this study was to determine the effects of supplemental leucine in milk replacer fed to neonatal lambs on pre- and postweaning growth, organ weight and serum amino acid profile. Twenty-four fall-born Dorset-cross twin or triplet ram lambs were weighed and allowed to nurse on dams for 12 hours to allow for colostrum intake. At 12 hours post-birth, lambs were assigned to one of two treatments: milk replacer (CON) and milk replacer with leucine added at 3% (LEU). Milk replacer was fed via a Lac-Tek milk machine. Lambs were provided milk replacer for ad libitum intake for 42 days. Lambs were weighed weekly until weaning. Ad libitum access to water, chopped hay and creep pellet were provided when lambs reached 2 weeks of age. On day 42, weaned lambs from both treatments were comingled in a common pen with access to creep and chopped hay. At approximately 50 pounds, lambs were sorted to individual pens for transition to a finishing diet. Feed was delivered daily and offered at 5% of lamb body weight to ensure ad libitum intake. Lambs were fed until one of two slaughter dates. The five heaviest lambs from each treatments were harvested after 67 days, and remaining lambs were harvested after 95 days. Leucine supplementation increased lamb weight during the pre-weaning phase from week two until week six (P < 0.001). Leucine during the milk-feeding phase had no effect on finishing weights, average daily gain (ADG) or days on feed. Lambs receiving leucine during the milk-feeding phase consumed more feed during the finishing phase than control lambs. Lambs did not differ in carcass characteristics except for body wall thickness (P = 0.05), in which leucine lambs had more fat. Our results indicate that leucine supplemented during the neonatal phase increases weaning weight. However, the differences in weight do not continue to finishing when fed for a common time on feed.
Introduction
The neonatal period is characterized by rapid skeletal muscle development, requiring sufficient amounts of amino acids (AA) to maximize growth. Furthermore, the neonatal phase allows for increased nutrient uptake as animals are milk-fed and milk bypasses the rumen.
Therefore, any milk ingested is available for digestion and absorption in the small intestine. The pre-ruminant stage allows for targeted nutrition to maximize animal growth.
Little is known about AA requirements in neonatal lambs; however, an imbalance of AA can lead to impaired growth and suppressed intake (Rogers and Egan, 1975). Previous data have shown that leucine is an essential AA that is involved in growth and development of muscle protein, although it is not an AA that is typically limiting in the diet.
Studies in neonatal piglets report that supplementing leucine can affect muscle growth positively and increase body weight, compared with control-fed animals (Sun et al., 2015; Boutry et al., 2016). However, leucine supplementation in lambs has been shown to result in no changes in growth but result in improved health (Mao et al., 2019).
The objective of this study was to determine the long-term effects of neonatal supplementation of leucine on growth during the pre- and postweaning phases, finishing performance and carcass characteristics.
Experimental Procedures
All procedures involving the use of animals were approved by the North Dakota State University (NDSU) Institutional Animal Care and Use Committee.
Nineteen neonatal fall-born Dorset-cross ram lambs (11.2 ± 0.33 pounds) were used in an experiment to determine long-term effects of neonatal supplementation of Leu in milk replacer. Lambs remained with ewes for 12 hours post-birth to ensure adequate colostrum intake and then were removed from ewes and sorted to a control milk replacer (Shepherd’s Choice, Premier1 Supplies, Washington, Iowa; CON, n = 10; Table 1) or milk replacer with leucine added at 3% (LEU, n = 9).
Lambs were trained to consume milk from a LAC-TEK Stainless 61450 (Biotic Industries Inc., Bell Buckle, Tenn.) milk dispenser that would allow for ad libitum milk intake. Both machines were calibrated to deliver the same amount of milk replacer and heated water.
Lambs were assigned randomly to a treatment and stratified by birth weight. Lambs were allowed access to milk for 42 days. Creep feed and chopped hay were provided for ad libitum intake when lambs reached 14 days of age (Table 2).
Table 1. Nutrient profile of milk replacer.
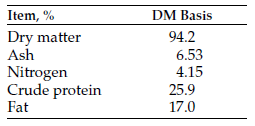
Table 2. Nutrient profile of feeds provided to lambs on a dry-matter basis.
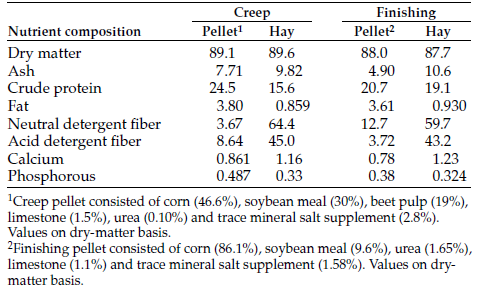
On day 42, lambs were removed from milk-feeding pens, comingled with lambs from both treatments, and provided ad libitum access to creep feed and chopped alfalfa hay. After all lambs were weaned, lambs then were moved to the NDSU Animal Nutrition and Physiology Center and group housed.
Lambs were moved to individual pens when they reached approximately 50 pounds to monitor daily feed intake. Creep feed and chopped alfalfa hay were provided at 5% of body weight (BW) and lambs then were transitioned to a finishing diet for 14 days. The finishing diet (Table 2) consisted of 90% pellet and 10% chopped hay.
Lambs were slaughtered at the NDSU Meat Laboratory on two days. The five heaviest lambs from each treatment were selected for slaughter after 67 days on feed, and all remaining lambs were slaughtered on the second day after 95 days on feed. After a 24-hour chill, carcasses were ribbed between the 12th and 13th rib to allow for carcass evaluation.
Lambs were weighed after birth and every seven days until weaning. Postweaning, lambs were weighed every 14 days until slaughter, and were weighed two consecutive days prior to slaughter. Samples of milk replacer, creep feed, chopped alfalfa hay and the finishing diet were sampled weekly. Carcasses were evaluated for fat depth, body wall thickness and ribeye area.
Nutrient profile of milk replacer, initial and final finishing weights, ADG, weight gain during the finishing phase and carcass characteristics were analyzed using the GLM procedure in SAS (SAS 9.4, SAS Institute Inc., Cary, N.C.). Lamb preweaning weights were analyzed using the MIXED procedure in SAS. Significance was declared at P ≤ 0.05.
Results and Discussion
Lambs receiving leucine-supplemented milk replacer weighed more than CON lambs from weeks two until six, when lambs were weaned (P < 0.001; Table 3). Neonatal effects of leucine did not carry over into the finishing phase, as evidenced by no differences in performance between treatments (Table 4). However, CON lambs consumed less feed than LEU lambs (P = 0.03).
Weight at slaughter tended to be different (P = 0.06), with LEU lambs weighing more than CON lambs (Table 5). However, weights of carcasses did not differ between treatments. Fat across the ribeye was not different between CON and LEU carcasses, and LEU carcasses had greater fat depth across the body wall than CON carcasses (P = 0.05).
Ribeye area also was not affected by treatment. Leg scores, flank streaking and overall carcass quality were not affected by leucine supplementation, and the percent of boneless, closely trimmed retail cuts (% BCTRC) also was not affected.
Table 3. Lamb weights during milk replacer feeding phase with or without supplemental leucine.
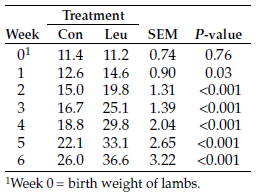
Table 4. Finishing performance of lambs fed milk replacer with or without supplemental leucine during the first 6 weeks of life.
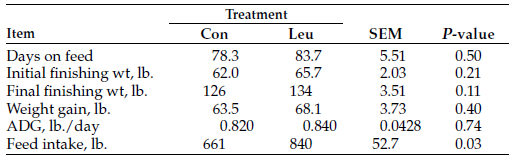
Table 5. Carcass characteristics of lambs fed milk replacer with or without supplemental leucine during the first 6 weeks of life.
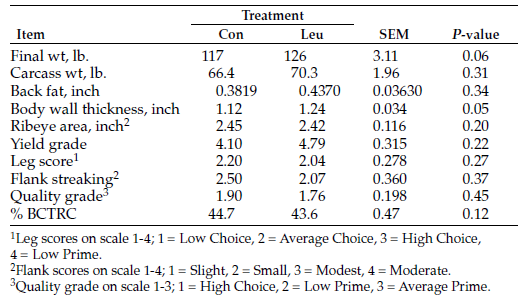
Lambs in this study were allowed ad libitum access to milk replacer with or without leucine. Leucine fed during this period may have been utilized by the body to increase muscle protein synthesis.
The preweaning growth performance of lambs in this study was different than the growth performance reported by Mao et al. (2019). Our lambs were allowed ad libitum access to milk replacer. Lambs in the study of Mao et al. were bottle fed three times daily, and daily allotments of leucine were delivered once daily (Mao et al., 2019). This individual delivery may have inhibited uptake and utilization of dietary leucine.
Further, the lambs were provided a mix of starter pellet and hay at the same time that allotment of milk replacer was being decreased (Mao et al., 2019). This shift in dietary availability, as well as its composition, may have influenced neonatal lamb growth.
During the finishing phase, CON lambs consumed less feed than LEU lambs and were slaughtered at similar body weights within a similar time frame. This may suggest that CON lambs experienced compensatory gain during the feeding phase, or were better able to digest and utilize nutrients from feed during this phase.
Lambs on the LEU treatment reached the benchmark weight for the transition to the finishing phase sooner than CON lambs. These lambs were transitioned to finishing diets earlier, and still fed to a common slaughter date. This extended period on feed may have contributed to the diminished performance of LEU lambs.
Mao et al. (2019) reported that control and leucine-supplemented lambs had similar growth performance and feed intake during the finishing phase. However, lambs from Mao et al. were a smaller-framed breed that would attain market weight earlier, and their finished weight is approximately half of the weight of the lambs used in this experiment (Mao et al., 2019).
In conclusion, supplemental Leu fed to milk-fed lambs increases early lamb performance and may increase producer profitability if lambs are sold at weaning. Additionally, Leu fed to young lambs does not have a detrimental effect on finishing or carcass performance of finished market lambs. Further evaluation of the effects of Leu on intestinal enzymes and muscle protein synthesis are warranted.
Acknowledgments
This project was supported by the North Dakota State Board of Agricultural Research and Education. The authors thank the employees of the NDSU Sheep Unit, Animal Nutrition and Physiology Center, the meat and nutrition laboratories at NDSU, and students who assisted with this project.
Literature Cited
Boutry, C.S. W. El-Kadi, A. Suryawan, J. Steinhoff-Wagner, B. Stoll, P.A. Orellana, H.V. Nguyen, S.R. Kimball, M.L. Fiorotto and T.A. Davis. 2016. Pulsatile delivery of a leucine supplement during long-term continuous enteral feeding enhances lean growth in term neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 310: E699-713.
Mao, H., C. Wang and Z. Yu. 2019. Dietary leucine supplementation enhances the health of early weaned Hu lambs. Anim. Feed Sci. Technol. 247:248-254.
Rogers, Q.R., and A.R. Egan. 1975. Amino acid imbalance in the liquid-fed lamb. Aust. J. Biol. Sci. 28:169-181.
Sun, Y., Z. Wu, W. Li, C. Zhang, K. Sun, Y. Ji, B. Wang, N. Jiao, B. He, W. Wang, Z. Dai and G. Wu. 2015. Dietary L-leucine supplementation enhances intestinal development in suckling piglets. Amino Acids. 47:1517-1525.
Male Fertility
Effects of feeding yearling beef bulls 60% dried distillers grains plus solubles or the equivalent as calcium sulfate on performance and reproductive attributes of yearling Angus bulls
Cierrah Kassetas1, Joel Caton1, James Kirsch1, Sheri Dorsam1, Matthew Crouse1, Kacie McCarthy1, Bryan Neville2, Kevin Sedivec3 and Carl Dahlen1
1Department of Animal Sciences, NDSU
2Carrington Research Extension Center, NDSU
3Central Grasslands Research Extension Center, NDSU
The objectives of this study were to determine the effects of dietary sulfur on semen quality and performance characteristics in yearling beef bulls. The results show no differences in initial body weight (BW), final BW, average daily gain (ADG), dry-matter intake (DMI), gain-to-feed (G:F) or scrotal circumference (SC). In sperm, the dried distillers grains plus solubles (DDGS)-treated bulls had decreased motile curvilinear velocity and motile velocity on an average path. Also, DDGS-treated bulls had increased progressive wobble and progressive linearity. The implications for this study are that sulfur may not be the only factor causing the observed differences in sperm characteristics.
Summary
Previous research in multiple species suggest that sulfur and/or dried distillers grains may influence semen quality in males. Thirty-six half-sibling beef bulls (13 months of age, BW = 485 kilograms [kg]) were assigned to one of three treatments: 1) corn-based diet containing 60% concentrate (CON, S = 0.2%, n = 12); 2) diet containing 60% DDGS as a replacement for corn (DDGS, S = 0.44% DM, n = 12); 3) CON diet+ equivalent sulfur of DDGS in the diet added as CaSO4 (SULF, S = 0.43%, n = 12). Diets were delivered to individual bulls in a Calan gate facility and fed to attain a targeted growth rate of 3.5 pounds per day during the course of a 112-day development period. Blood was collected for analysis on day 0, 56, and 112. Scrotal circumference and semen were collected on day 0, 28, 56, 84 and 112. Semen was collected using a Pulsator IV electronic ejaculator (Lane Manufacturing Inc., Denver, Colo.). Sperm concentration, motility and morphology characteristics were assessed using Computer Assisted Semen Analysis (IVOS II, Hamilton Thorne, Beverly, Mass.) to determine the overall semen concentration, percent motile, percent morphology and percent progressive. By design, no differences (P = 0.14) were observed among treatments for ADG or final BW. In addition, no differences (P = 0.14) were observed among treatments for DMI or scrotal circumference. However, bulls in the SULF treatment tended (P = 0.07) to have reduced feed conversion, compared with bulls in the CON treatment. No differences (P = 0.18) were observed among treatments in ejaculate characteristics including volume, sperm concentration or proportion of sperm being classified as motile, progressively motile, slow or static. However, in sperm classified as motile, the average path velocity (VAP) and curvilinear velocity (VCL) were greater (P ≤ 0.03) for CON bulls, compared with DDGS bulls, whereas SULF were intermediate. In sperm classified as progressive, wobble and linear were greater (P = 0.004) for DDGS bulls when compared with CON bulls, whereas SULF were intermediate. We found no difference (P ≥ 0.13) among treatments for concentrations of testosterone (T) or thyroxine (T4) in serum; however, a decrease (P = 0.009) occurred in triiodothyroxine (T3) concentrations for DDGS bulls. In yearling bulls that were managed to attain similar growth rates, the addition of sulfur tended to reduce feed conversion but did not impact hormone or sperm characteristics. However, although not influencing measures of semen quality traditionally used in breeding soundness evaluations, feeding 60% DDGS in the diet of developing bulls reduced concentrations of T3 and altered semen kinematics. Therefore, the influence of DDGS on hormone and sperm characteristics is likely because of factors other than excess dietary sulfur.
Introduction
Dried distillers grains plus solubles has a three-fold greater concentration of protein, fat, fiber and phosphorus, when compared with corn. In addition, sulfuric acid is added to maintain pH levels which results in DDGS having a greater concentration of sulfur that can range from 0.33% to 0.74% of the diet (U.S. Grains Council, 2016; Buckner et al., 2011). The maximum tolerable concentration of sulfur is recommended to be less than 0.4% of the diet; however, depending on the diet composition, this percentage could change (NASEM, 2016).
Studies in ram lambs show that increased sulfur levels from DDGS can decrease testosterone levels and increase sperm abnormalities (Van Emon et al., 2013; Crane et al., 2015). In addition, studies in male mice show a decrease in sperm motility and morphology when sulfur dioxide (SO2) was released into ambient air (Meng et al. 2004; Zhang et al.; 2016a).
However, in developing bulls no research has been done looking at the effect of DDGS on reproduction. The increase of sulfur from DDGS may be impacting semen quality and quantity. This study will look at how two different sources of dietary sulfur affect semen quality and feed efficiency in yearling beef bulls.
Experimental Procedures
All procedures were approved by the North Dakota State Institution for Animal Care and Use Committee.
Thirty-six half- sibling beef bulls (13 months of age, BW = 485 kg) were assigned to one of three treatments (Table 1): 1) corn-based diet containing 60% concentrate (CON, S = 0.2%, n = 12); 2) diet containing 60% DDGS as a replacement for corn (DDGS, S = 0.44% DM, n = 12); or 3) CON diet+ equivalent sulfur of DDGS in diet added as CaSO4 (SULF, S = 0.43%, n = 12). Bulls were fed individually in Calan gates, and diets were adjusted to target a 3.5 pounds per day ADG.
Table 1. Diet composition and % DM inclusion for the three treatments.
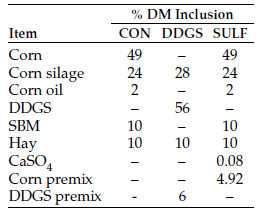
Blood was collected for analysis on days 0, 56 and 112. Scrotal circumference (SC) and semen were collected on days 0, 28, 56, 84 and 112. For SC measurements, a scrotal tape was placed around the widest portion of the scrotum. Semen was collected using a Pulsator IV electronic ejaculator (Lane Manufacturing Inc., Denver, Colo.) and analyzed using a Computer Assisted Semen Analysis (CASA, IVOS II, Hamilton Thorne, Beverly, Mass.) instrument.
The overall semen concentration, percent motile, percent morphology and percent progressive of semen were determined. In addition, kinematic parameters, including average path velocity (VAP), curvilinear velocity (VCL), velocity on a straight line (VSL), linear and wobble were evaluated for the motile and progressive populations of sperm. Puberty was defined when bulls had a scrotal circumference (SC) equal to or greater than 30 centimeters (cm), equal to or greater than 30% motile sperm, and equal to or greater than 70% normal sperm.
Blood plasma was collected using vacutainer tubes, and an Immulite 1000 (Siemens Healthineers, Erlangen, Germany) was used to determine testosterone (T), triiodothyroxine (T3) and thyroxine (T4) concentrations in serum. Data were analyzed using Statistical Analysis System (SAS Inst. Inc.; Cary, N.C.) and P-values ≤ 0.05 were considered significant. Performance data were analyzed using the GLM procedure in SAS. Semen characteristics data were analyzed using the repeated measures analysis in the MIXED procedure with body weight set as a covariate.
Results and Discussion
By design no differences (P ≥ 0.14) were observed among treatments for ADG and final BW (Table 2). Bulls developed at faster rates of gain have greater scrotal circumference and enhanced semen characteristics compared with slower gaining counterparts (Palasz et al., 1994).
Table 2. The effect of dietary sulfur from DDGS or CaSO4 on beef bull performance characteristics in beef bulls.
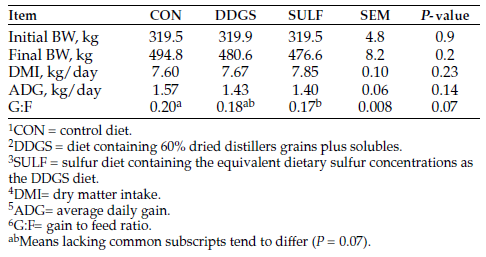
However, growth rates do have an upper limit as bulls managed very aggressively and gaining greater than 3.8 pounds/day had reduced semen parameters compared with bulls managed to gain equal to or less than 2.2 pounds/day (Skinner et al., 1981). Because the optimal growth rates for yearling bulls was found to be between 2.2 and 3.5 pounds/day (Brito et al., 2012) bulls in all treatments of the current report were managed to have a targeted average daily gain (ADG) of 3.5 pounds/day to ensure that any differences observed were due to dietary treatment composition and not to rate of gain.
We observed no differences in DMI, but feed conversion tended to be reduced (P = 0.07) in SULF bulls compared with CON (Table 2). Previous research suggests that steers fed ad libitum diets containing 0.46% dietary sulfur caused a reduction in ADG and DMI with a tendency to reduce feed conversion when compared with the control diet containing 0.13% sulfur (Spears et al. 2011).
Neville et al. (2012) also found that for bulls fed ad libitum diets containing 20% (0.7% S), 40% (0.7% S) or 60% (0.9% S) DDGS (DM basis) had a linear decrease in ADG and DMI with a tendency to decrease feed conversion. In the studies from Spears et al. (2011) and Neville et al. (2012), tendencies for a decrease in feed efficiency was observed in steers fed for ad libitum intake whereas, in the current study, a tendency to decrease feed efficiency was also observed in bulls that were fed to target 3.5 pounds/day.
These bulls began the project at 13 months of age where only 22% were considered pubertal. As the bulls aged and body weight increased the proportion of bulls that were pubertal increased (Figure 1) but was not affected by treatments (P = 0.36). Interestingly, all bulls were considered pubertal by 13 months of age (day 112).
Figure 1. Percent of bulls that were pubertal during the five collections.
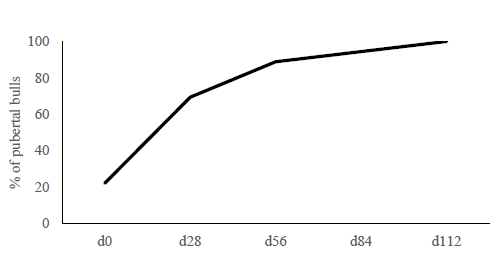
No differences were observed for measures evaluated in traditional breeding soundness examinations: scrotal circumference, ejaculate volume, sperm concentration, percent motile or progressively normal and percent with normal morphology (Table 3).
To analyze sperm kinematics on the CASA machine individual sperm are placed into categories of motile, progressive and slow based on their motion characteristics, then data are analyzed for sperm within each respective category. For sperm in the motile category, curvilinear velocity (VCL) was reduced in the DDGS treatment (P = 0.03), compared with the CON, whereas SULF was intermediate. In addition, velocity along an average path (VAP) was reduced (P = 0.02) in DDGS bulls, compared with CON bulls (Table 4).
Table 3. Overall semen characteristics.
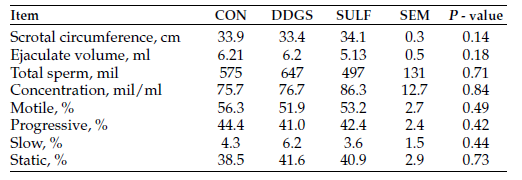
Table 4. CASA motion parameter measurements for sperm.
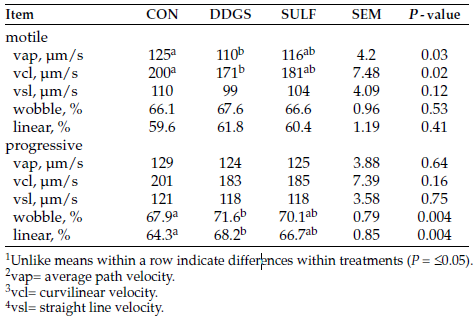
Table 5. Hormone levels in the blood.
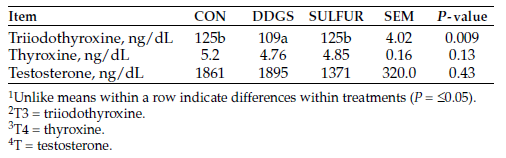
A reduction in both of these traits indicates that sperm in the DDGS-treated bulls may be moving slower. The sperm in the progressive category, wobble and linear, were greater in DDGS bulls (P = 0.004 and P = 0.004, respectively), when compared with CON, where SULF was intermediate. The increase in progressive wobble may indicate an increase in oscillation of the sperm head for the DDGS bulls. An increase in progressively linear sperm suggests that the sperm had a decreased amplitude for curvilinear path and direction.
We found no differences (P ≥ 0.13) among treatments for concentrations of testosterone (T) or thyroxine (T4) concentration in the serum. However, we observed a decrease (P = 0.009) in triiodothyroxine (T3) concentrations for bulls fed DDGS (Table 5).
One potential mechanism for the decrease in T3 concentrations could be the relationship between sulfur and selenium. With the increase of ruminal sulfur, selenium is reduced which reduces T3 concentrations (Ivancic and Weiss, 2001; Hosnedlova et al., 2017).
This decrease of T3 could be decreasing overall selenium levels in DDGS-treated bulls. This could lead to a decrease in glutathione peroxidase activity and nonspecific immune cell response (Spears, 2000), resulting in cells becoming more susceptible to oxidative damage (Hosnedlova et al., 2017). Oxidative damage within the testes could be one explanation for the altered sperm kinematics in the DDGS-treated bulls.
In yearling bulls that were managed to attain similar growth rates the addition of sulfur tended to reduce gain:feed but did not impact hormone or sperm characteristics. However, although not impacting measures of semen quality traditionally used in breeding soundness evaluations, feeding 60% DDGS in the diet of developing bulls reduced concentrations of T3 and altered semen kinematics.
Therefore, the impact that DDGS has on hormone and sperm characteristics is not exclusively due to excess dietary sulfur in DDGS. Further research is necessary to elucidate what factors within DDGS are impacting semen quality, and whether effects observed are influencing sperm fertilization potential.
Acknowledgments
The authors thank the North Dakota Corn Council and the North Dakota Agriculture Experiment Station for the funding for this project. Also, we thank the staff at the Animal Nutrition and Physiology Center for the assistance in management of the bulls.
Literature Cited
Brito, L.F.C., A. D. Barth, R.E. Wilde and J.P. Kastelic. 2011. Effect of growth from 6 to 16 months of age on sexual development and reproductive function in beef bulls. Theriogenology 77: 1398- 1405.
Buckner, C.D., M.F. Wilken, J.R. Benton, S.J. Vanness, V.R. Bremer. 2011. Nutrient variability for distillers grains plus solubles and dry matter determination of ethanol by-products. Prof. Anim. Sci. 27:57–64.
Hosnedlova, B., M. Kepinska, S. Skalickova, C. Fernandez, B. Ruttkay-Nedecky, T.D. Malevu, M. Baron, M. Melcova, J. Zidkova, R. Kizek. 2017. A summary of new findings on the biological effects of selenium in selected animal species- a review. Int. J. Mol. Sci. 18:2209.
Meng, Z., and W. Bai. 2004. Oxidation damage of sulfur dioxide on testicles of mice. Environ. Research 96: 298-304.
National Academics of Sciences, Engineering, and Medicine. 2016. Nutrient Requirements for Beef Cattle, 8th rev. ed. Washington, D.C.: The National Academies Press.
Neville, B.W., G.P. Lardy, K.K. Karges, S.R. Eckerman, P.T. Berg and C.S. Schauer. 2012. Interaction of corn processing and distillers dried grains with solubles on health and performance of steers. J. Anim. Sci. 90:560–567.
Palasz, A.T., W.F. Cates, A.D. Barth and R.J. Mapletoft. 1994. The relationship between scrotal circumference and quantitative testicular traits in yearling beef bulls. Theriogenology 42: 715-726.
Skinner, J.D. Nutrition and fertility in pedigree bulls. In: Environmental Factors in Mammalian Reproduction, Gilmore D, Cook B (Eds.), MacMillan Publishers, 1981, 160-168.
Spears, J.W. 2000. Micronutrients and immune function in cattle. Proceedings of the Nutrition Society 59: 587-594.
Spears, J.W., K.E. Lloyd and R.S. Fry. 2011. Tolerance to cattle increased dietary sulfur and effect of dietary cation- anion balance. J. Anim. Sci. 89:2502-2509.
U. S. Grains Council. 2016. A Guide to Distiller’s Dried With Solubles, 3rd ed. Washington, D.C.
Zhang, J.H., L. Zhihui, Q. Mingli, R. Zheng, J. Shetty and J. Wang. 2016. Sodium fluoride and sulfur dioxide affected male reproduction by disturbing blood-testis barrier in mice. Food and Chem. Toxicol. 94: 103-111.
Effects of feeding 60% dried distillers grains plus solubles or the equivalent sulfur as calcium sulfate on hydrogen sulfide gas production in the rumen of yearling Angus bulls
Cierrah Kassetas1, Bryan Neville2, Joel Caton1, James Kirsch1, Kacie McCarthy1, Kevin Sedivec3, Matthew Crouse1 and Carl Dahlen1
1Department of Animal Sciences, NDSU
2Carrington Research Extension Center, NDSU
3Central Grasslands Research Extension Center, NDSU
The objective of this study was to evaluate the influence of feeding 60% dried distillers grains plus solubles (DDGS) or the equivalent sulfur from calcium sulfate (CaSO4) to yearling beef bulls on ruminal hydrogen sulfide (H2S) concentrations. This study found increasing levels of hydrogen sulfide in the rumen when greater levels of sulfur are fed as DDGS or as calcium sulfate. This study indicates that an increased percent of sulfur in the diet will lead to increases in ruminal H2S concentrations in yearling beef bulls.
Summary
Twenty-four crossbred beef bulls (13 months of age, BW = 320 kilograms [kg]) were assigned one of three treatments: 1) corn-based diet containing 60% concentrate (CON, S = 0.2%, n = 8); 2) diet containing 60% DDGS as a replacement for corn (DDGS, S = 0.44% DM, n = 8); 3) CON diet + equivalent sulfur of DDGS in diet added as CaSO4 (SULF, S = 0.43%, n = 8). Ruminal gas samples were taken via rumen puncture four to six hours post feeding on days zero, 14 and 42. Concentrations of H2S were determined using hydrogen sulfide detector tubes, which were connected to a volumetric gas pump (Gastec, Kanawaga, Japan). A day × treatment interaction was observed for ruminal H2S (P = 0.005). No difference (P = 0 .06) was observed in bulls fed DDGS or SULF on H2S concentrations on day 14, while animals fed CON had lower H2S concentration (P = 0.004). On day 42, concentrations of H2S were greater in SULF than all other treatments (P ≤ 0.006), while DDGS was greater than CON (P ≤ 0.05). Elevated H2S in bulls fed the SULF treatment, compared with the DDGS treatment, may indicate that ruminal availability of sulfur (S) in CaSO4 was greater than the S in DDGS.
Introduction
Dried distillers grains plus solubles (DDGS) is a co-product of the ethanol industry that has been used increasingly in beef cattle diets. Dried distillers grains plus solubles is made from the conversion of corn to ethanol.
There is a three-fold increase in the amount of protein, fat, fiber and phosphorus when corn is converted to DDGS. After the dry grinding process, sulfuric acid is added to slow fermentation while maintaining pH levels to convert starch to ethanol. This addition of sulfuric acid increases the sulfur concentration of this co-product (U.S. Grains Council, 2016).
A study by Neville et al. (2012) found a decrease in average daily gain (ADG) and dry-matter intake (DMI) as the concentration of DDGS increased. A paper by Drewnoski et al. (2014) discussed the impacts of sulfur from DDGS and non-DDGS sources on beef cattle performance. They found that DMI and ADG decreases with the increase of DDGS in the diet, indicating that excess sulfur from non-DDGS sources also may impact the animal performance.
In more serious cases, polioencephalomalacia (PEM) can occur. Polioencephalomalacia is a neurological disease in ruminants that is a result of sulfur toxicity. Therefore, the hypothesis for this study was that bulls consuming elevated concentrations of sulfur as sulfuric acid from DDGS or a non-DDGS source as CaSO4 will increase ruminal H2S.
The objective of this study was to evaluate the influence of feeding 60% dried distillers grains plus solubles (DDGS) or the equivalent sulfur from calcium sulfate (CaSO4) to yearling beef bulls on ruminal hydrogen sulfide (H2S )concentrations.
Experimental Procedures
All procedures were approved by the North Dakota State Institution for Animal Care and Use Committee.
Twenty-four crossbred beef bulls (13 months of age, BW = 320 kilograms [kg]) were assigned one of three treatments: 1) corn-based diet containing 60% concentrate (CON, S = 0.2%, n = 8); 2) diet containing 60% DDGS as a replacement for corn (DDGS, S = 0.44% DM, n = 8); 3) CON diet + equivalent sulfur of DDGS in diet added as CaSO4 (SULF, S = 0.43%, n = 8). Bulls were individually fed in Calan gates to target a 3.5 pound/day ADG.
Ruminal gas samples were taken via rumen puncture four to six hours post feeding on days zero, 14 and 42. Landmarks to determine the site for rumen puncture were the midpoint between the last rib and spine.
The site was clipped and prepared by alternating isopropyl alcohol and Betadine scrubs three times. After the samples were obtained, the needle was removed and a 10% iodine solution was sprayed on the sampling site. Concentrations of H2S were determined using hydrogen sulfide detector tubes which were connected to a volumetric gas pump (Figure 1, Gastec, Kanawaga, Japan).
Data were analyzed as repeated measures using the MIXED procedures of SAS for effects of day, treatment, and a day × treatment interaction. Differences were considered significant at a P- value ≤ 0.05.
Figure 1. H2S Sampling apparatus.
Results and Discussion
Bulls were managed to have a consistent ADG to ensure that differences existed because of an effect of treatment rather than an increase in body weight. Therefore, by design, no differences were observed for initial body weight (BW), final BW, ADG or gain:feed (G:F) ratio (P = 0.10, 0.49, 0.14, and 0.21, respectively).
However, bulls consuming DDGS had reduced dry-matter intake (P = 0.04), compared with bulls in the CON or SULF treatments. Dry matter intake was reduced in DDGS bulls because these bulls were delivered less feed when compared with other treatments to maintain the targeted ADG of 3.5 pounds/day.
In Figure 2, a day × treatment interaction was observed for ruminal H2S (P = 0.005). On day 0, there was no effect of treatment on ruminal H2S concentration (P = 0.97).
No differences (P = 0.06) were observed on day 14 when bulls were fed DDGS or SULF, while animals fed CON had lower ruminal H2S concentrations (P = 0.004). On day 42, DDGS and SULF increased ruminal H2S concentrations (P ≤ 0.006). Furthermore, SULF increased ruminal H2S concentrations greater than CON or DDGS on day 42 (P ≤ 0.05).
Elevated ruminal H2S in bulls fed the SULF diet, compared with DDGS, may indicate that ruminal availability of sulfur (S) in CaSO4 was greater than that of sulfur in DDGS. Overall, an increased percent of sulfur in the diet will lead to increase in ruminal H2S in yearling beef bulls.
Table 1. Diet composition and % DM inclusion for the three treatments.
Table 2. Effects of dietary sulfur on yearling beef bull performance characteristics.
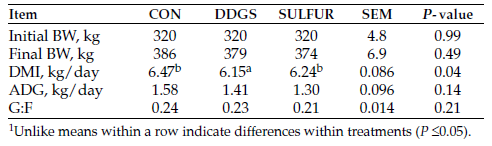
Acknowledgments
The authors thank the North Dakota Corn Council and the North Dakota Agriculture Experiment Station for the funding for this project. Also, we thank the staff at the Animal Nutrition and Physiology Center for the assistance in management of these animals.
Literature Cited
Drewnoski, M.E., D.J. Pogge and S.L. Hansen. 2014. High sulfur in beef cattle diets: a review. J. Anim. Sci. 92: 3763- 3780.
Neville, B.W., G.P. Lardy, K.K. Karges, S.R. Eckerman, P.T. Berg and C.S. Schauer. 2012. Interaction of corn processing and distillers dried grains with solubles on health and performance of steers. J. Anim. Sci. 90:560–567.
U. S. Grains Council. 2016. A Guide to Distiller’s Dried With Solubles (3rd ed.). DDGS User Handbook. Washington, D.C.
Influences of dietary flax on male reproductive traits when supplemented prior to breeding in sheep
Paige P. Anderson1, Amanda M. Long1, Ethan R. Schlegel2, Travis W. Hoffman3, Kimberly A. Vonnahme3,4, Jennifer L. Momsen5, Lauren L. Hulsman Hanna3, James D. Kirsch3, Carl R. Dahlen3, Kacie L. McCarthy3 and Christopher S. Schauer1
1Hettinger Research Extension Center, NDSU
2New Generation Supplements, Belle Fourche, S.D.
3Department of Animal Sciences, NDSU
4Zoetis, Parsippany, N.J.
5Department of Biological Sciences, NDSU
The objective of the present study was to examine the influence of flaxseed supplementation on male reproductive traits during a 112-day period leading up to the breeding season. Results indicate that Flaxlic Sheep Tubs did not affect male reproductive traits when supplemented to a basal diet containing soybean hulls, corn and soybean meal.
Summary
Fertility in rams is of critical importance in range sheep operations. Prior to breeding, fertility can be enhanced by improving nutritional management. Fatty acid (FA) supplementation has been shown to improve male reproductive characteristics, such as sperm motility, concentration and morphology. Supplementation with flax prior to breeding is a potential strategy to increase omega-3 FAs in the diet. The objective of this study was to evaluate the effectiveness of flax supplementation on serum testosterone concentration and semen quality. One hundred twenty Rambouillet ram lambs (92.4 ± 6.12 pounds) were assigned randomly to 24 pens (five rams/pen; n = 12) and fed for 112 days. Rams were assigned to receive a Flaxlic® Sheep Tub (FLX) or a control (CON). Two-day weights were taken on days minus 1 and 0 and days 111 and 112. Serum for testosterone concentration analysis, semen for quality analysis and scrotal circumference measurements were collected on days 83 and 84 and 111 and 112. Average daily gain (ADG) for FLX rams was not different from CON (P = 0.25). No differences were observed for testosterone concentrations between CON and FLX treatments (P = 0.70). We found no differences in scrotal circumference, sperm motility, sperm morphology or sperm concentration (P ≥ 0.15). These results are in contrast to other research in which flax was fed to rams. Therefore, flaxseed supplementation in a tub form is not recommended to growing ram lambs for the improvement of semen quality when the basal diet consists of soybean hulls, corn and soybean meal.
Introduction
Reproduction is a vital component for any range sheep operation. The male side of reproduction is a component of overall productivity that can be overlooked.
One way to enhance a ram’s performance during the breeding season is to improve the nutrients in the ram’s diet leading up to breeding. Not only will this help rams regain lost condition from the previous breeding season, but it may stimulate improved spermatogenesis and sperm cell function. Adding extra nutrients to the pre-breeding ration, such as essential fatty acids, has been shown to further improve a male’s reproductive efficiency (Baiomy and Mottelib, 2009; Yan et al., 2013; Esmaeili et al., 2014).
Flaxseed provides two essential fatty acids (FAs): Alpha-linolenic acid (ALA; C18:3 ω-3), an omega-3 (ω-3) FA and linoleic acid (LA: C18:2 ω-6), an omega-6 (ω-6) FA. Flax is approximately 45% oil. Of the total fats in flax oil, about 57% is ALA and 16% percent is LA (Morris, 2007).
Flaxseed in particular has been shown to improve sperm motility and progressive motility in bulls (Moallem et al., 2015). Flaxseed also has been shown to increase levels of the reproductive hormones such as the gonadotropin-releasing hormone, follicle stimulating hormone (Yan et al., 2013), luteinizing hormone (Yan et al., 2013) and testosterone (Baiomy and Mottelib., 2009; Yan et al., 2013; Esmaeili et al., 2014). Supplementing male sheep with flaxseed prior to breeding may be a way to improve semen quality and thereby improve fertility.
Our hypothesis was that the supplementation of flaxseed in the diet for rams would increase testosterone in the blood and therefore improve spermatogenesis and reproductive performance while preventing sperm abnormalities. The objective of the present study was to supplement flaxseed in an applied setting using Flaxlic® Sheep Tubs during a 112-day period leading up to the breeding season.
Experimental Procedures
Rambouillet ram lambs (n = 120) were selected from the NDSU Hettinger Research Extension Center flock. Ram lambs (approximately 4 months of age; 92.4 ± 6.12 pounds) were assigned randomly to 24 pens (five rams/pen; 271.25 feet2/ram), with pen serving as the experimental unit. Rams were fed a basal ration with a Flaxlic® Sheep Tub (FLX; n = 12) or a control ration alone (CON; n = 12). The basal ration was a total mixed ration (TMR) made up of 60% soybean hulls, 10% corn, 15% soybean meal and 15% Market Lamb Supplement (dry matter basis; Southwest Feed Inc.).
Two-day weights and body condition score were taken on days minus 1 and 0 and days 111 and 112, with ram body weight recorded every 28 days to monitor ram health. Ram scrotal circumferences were taken during a two-day period alongside semen collection on days 83 and 84, and again on days 111 and 112.
Blood and semen were collected during a two-day period on days 83 and 84, then again on days 111 and 112. Blood was collected via jugular venipuncture for testosterone analysis. Semen was collected via electro-ejaculation during a two-day period on days 83 and 84, then again on days 111 and 112.
The first successful ejaculate from each ram was evaluated. Sperm morphology and motility traits were analyzed for reproductive efficiency.
Results and Discussion
Performance
Initial and final ram weight or body condition scores (BCS) did not differ between treatments (Table 1; P ≥ 0.25). As the rams were given free access to feed with or without the addition of the Flaxlic® Sheep Tub, this result was expected.
However, we found a day effect for weight gain (P < 0.001), which also was expected due to the growth of a ram lamb into a mature ram. We observed a treatment × day interaction (P = 0.04) for average daily gain (ADG). Between days 28 and 56, the CON treatment had a higher ADG than the FLX group (P = 0.04). The CON treatment gained more through time than the FLX group.
We observed no effect of treatment on daily dry matter intake between CON and FLX treatments (P = 0.23). Intake of the Flaxlic® Sheep Tub was 0.09 pound per head-1 per day1, or 2.2% of their total feed intake per day. The gain-to-feed (G:F) ratio was not different between CON and FLX treatments (P = 0.77)
We observed no treatment × day interactions for scrotal circumference (SC; P = 0.34). We found a day effect between days 83 and 84 and days 111 and 112 (31.92 and 34.30 centimeters, respectively; P < 0.001), as expected of maturing ram lambs (Camela et al., 2018). We also found no treatment effects for overall scrotal circumference (SC) (P = 0.72) or on days 83 and 84 (Table 2; P = 0.56).
Table 1. Influence of Flaxlic® Sheep Tubs on weight and body measurements in Rambouillet ram lambs.
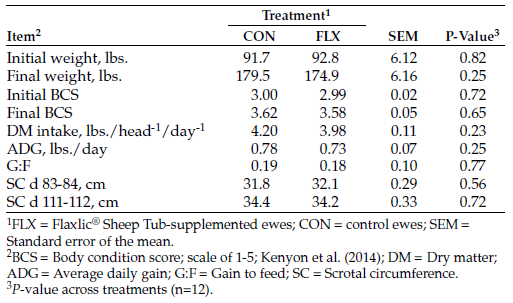
Table 2. Influence of Flaxlic® Sheep Tubs supplementation on semen characteristics in Rambouillet ram lambs.
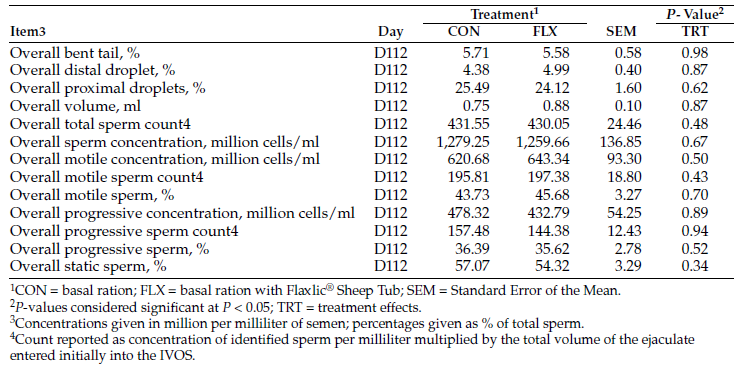
The present study used a less protected form of flax in the tub, made up of flax oil and flaxseed meal. Changes in scrotal circumference are affected by maturity and season. Rams were in the correct season to stimulate changes in SC. Therefore, the lack of change in SC simply may be due to the rams’ immaturity.
Testosterone
We observed no treatment × day interactions (P = 0.99) for serum testosterone concentrations. Serum testosterone was not different between treatments (P = 0.99). A day effect was observed between days 83 and 84 and 111 and 112 (P = 0.02). Testosterone increased through time, which is in unison with the observed increase in scrotal circumference.
Sperm Morphological Abnormalities
We found no treatment × day interactions (P ≥ 0.08) or treatment effects (P ≥ 0.62) for sperm abnormalities (Table 2). The percentage of bent tails and total abnormalities were affected by day (P ≤ 0.04). The bent tail percentage and total abnormalities decreased as rams aged, which was expected due to the age of the rams.
The rams in the present study were fed processed flax oil and flaxseed meal in the form of a tub, in addition to the high levels of omega-6 FAs found in the basal ration. We found no treatment x day interactions (P ≥ 0.15) for semen volume or concentration.
We also observed no effects of treatment on semen volume, total count or concentration (P ≥ 0.48). We did find a day effect (P ≤ 0.03) for ejaculate volume, sperm concentration and total sperm count, with all variables increasing as the rams aged.
Motility
We found no treatment × day effects (P ≥ 0.18) for any motility measurement. We also observed no effect of treatment (P ≥ 0.42) for any motility measurement. We found a day effect (P = 0.05) for the progressive sperm count, decreasing from 184.85 sperm per milliliter (ml) on days 83 and 84 to 150.75 sperm per ml on days 111 and 112. We are unsure why the progressive sperm count decreased because this value should have increased as the rams aged.
Results and Discussion
These results are in contrast with Pesta and Drouillard (2010), who reported increased ADG and improved feed efficiency between treatments in Flaxlic® Tub-fed bulls (P < 0.05). Pesta and Drouillard (2010) conversely reported the control group of bulls had higher feed intakes than Flaxlic® Tub-supplemented bulls. Pesta and Drouillard (2010) also reported improved G:F ratios not found in the present study.
The present study results were opposite to the similar study in bulls, possibly indicating a difference in physiological response to flax between bulls and rams. This may be due to a species-specific response that does not occur in sheep. Another explanation may be the rams were not eating enough of the Flaxlic® Sheep Tub, compared with the intake of the bull. Rambouillet rams may require more flax per pound of bodyweight than bulls.
The lack of change in SC is in agreement with Baiomy and Mottelib (2009), who also found no change in SC between flaxseed-supplemented and control rams, reporting an increase of testosterone in the flax-fed rams versus the control after two months of treatment. These results are in agreement with Pesta and Drouillard (2010), who reported no differences between control and Flaxlic® supplemented bulls for the percent of normal sperm and decreased abnormal sperm in rams supplemented with unprotected flax oil. The addition of a Flaxlic® Sheep Tub did not improve reproductive parameters or influence testosterone concentration level.
Acknowledgments
The authors thank New Generation Feeds and AmeriFlax for funding this research.
Literature Cited
Baiomy, A.A., and A.A. Mottelib. 2009. Effect of dietary flaxseed oil supplementation on reproductive performance of rams during summer. Anim. Hyg. Sust. Livest. Prod. 3:1263–1265.
Camela, E.S.C., R.P. Nociti, V.J.C. Santos, B.I. Macente, M. Murawski, W.R.R. Vicente, P.M. Bartlewski and M.E.F. Oliveira. 2018. Changes in testicular size, echotexture, and arterial blood flow associated with the attainment of puberty in Dorper rams raised in a subtropical climate. Reproduction in Domestic Animals.
Esmaeili, V., A.H. Shahverdi, A.R. Alizadeh, H. Alipour and M. Chehrazi. 2014. Saturated, omega-6 and omega-3 dietary fatty acid effects on the characteristics of fresh, frozen-thawed semen and blood parameters in rams. Andrologia. 46:42–49.
Moallem, U., N. Neta, Y. Zeron, M. Zachut and Z. Roth. 2015. Dietary α-linolenic acid from flaxseed oil or eicosapentaenoic and docosahexaenoic acids from fish oil differentially alter fatty acid composition and characteristics of fresh and frozen-thawed bull semen. Theriogenology. 83:1110–1120. doi:10.1016/j.theriogenology.2014.12.008.
Morris, D. 2007. Flax - A Health and Nutrition Primer. 4th ed. Flax Council of Canada. Available from: https://flaxcouncil.ca/resources/nutrition/technical-nutrition-information/flax-a-health-and-nutrition-primer.
Pesta, A.C., and J.S. Drouillard. 2010. FlaxLic supplementation improves growth performance of Angus bulls. Kansas Agricultural Experiment Station Research Reports. 108–110.
Yan, L., X. Bai, Z. Fang, L. Che, S. Xu and D. Wu. 2013. Effect of different dietary omega-3/omega-6 fatty acid ratios on reproduction in male rats. Lipids in Health and Disease. 12:33.

(Photo by Sarah Underdahl, NDSU)
Backgrounding / Finishing
Discovering value in North Dakota calves: Dakota Feeder Calf Show feedout project XVII 2018-2019
Karl Hoppe1 and Dakota Feeder Calf Show Livestock Committee2
1Carrington Research Extension Center, NDSU
2Turtle Lake, N.D.
North Dakota cattle producers are identifying cattle with superior growth and carcass characteristics by participating in the Dakota Feeder Calf Show. Average profitability between consignments from the top five herds and the bottom five herds was $169.47 per head for the 2018-2019 feeding period.
Summary
The Dakota Feeder Calf Show feedout project helps North Dakota cattle producers discover the actual value of their spring-born beef steer calves, provide comparisons among herds, and benchmark feeding and carcass performance. Cattle consigned to the feedout project were delivered to the Carrington Research Extension Center Livestock Unit on Oct. 20, 2018. After a 212-day feeding period with 2.33 percent death loss, cattle averaged 1,325.4 pounds (shrunk harvest weight). Feed required per pound of gain was 7.4 (dry-matter basis). Overall pen average daily gain was 3.24 pounds. Feed cost per pound of gain was $0.487 and total cost per pound of gain was $0.746. Profit ranged from $142.46 between the top 5 herds and bottom 5 herds per head for pen-of-three cattle with superior growth and carcass traits to minus $27.01 per head (no death loss). Substantial variability in the feeding and carcass value of spring-born calves continues to be discovered through participation in the feedout project.
Introduction
Cow-calf producers need to remain competitive with other livestock and poultry in the meat industry. By determining calf value in a feedout program, cow-calf producers can identify superior genetics under common feedlot management. Marketplace premiums are provided for calves that have exceptional finishing performance and produce a high-quality carcass.
Cost-effective feeding performance is needed to justify the expense of feeding cattle past weaning. Because North Dakota has low-cost feeds and a favorable climate, low cost per pound of gain can be accomplished (Hoppe et al., 1997).
Combining the low cost of gains with the identification of superior cattle, this ongoing finishing project provides cattle producers with an understanding of cattle feeding and cattle selection in North Dakota.
Experimental Procedures
The Dakota Feeder Calf Show was developed for cattle producers willing to consign steer calves to a show and feedout project. The calves were received in groups of three or four on Oct. 20, 2018, at the Turtle Lake Weighing Station, Turtle Lake, N.D., for weighing, tagging, veterinary processing and showing. The calves were evaluated for conformation and uniformity, with the judges providing a discussion to the owners at the beginning of the feedout. The number of cattle consigned was 129, of which 110 competed in the pen-of-three contest.
The calves then were shipped to the Carrington Research Extension Center, Carrington, N.D., for feeding. Prior to shipment, calves were vaccinated, implanted with Synovex-S, dewormed and injected with a prophylactic long-acting antibiotic.
Calves then were sorted and placed on corn-based receiving diets. After an eight-week backgrounding period, the calves were transitioned to a 0.62 megacalorie of net energy for gain (Mcal NEg) per pound finishing diet. Cattle were weighed every 28 days, and updated performance reports were provided to the owners. Cattle were reimplanted with Synovex-Plus.
An open house was held on Feb. 1, 2019, at the Carrington Research Extension Center Livestock Unit, where the owners reviewed the calves and discussed marketing conditions.
The cattle (125 head) were harvested on May 22, 2019. The cattle were sold to Tyson Fresh Meats, Dakota City, Neb., on a grid basis, with premiums and discounts based on carcass quality. Carcass data were collected after harvest.
Ranking in the pen-of-three competition was based on the best overall score. The overall score was determined by adding the index values for finishing average daily gain (25% of score), marbling score (25% of score) and profit (25% of score) and subtracting index value for calculated yield grade (25% of score). The Dakota Feeder Calf Show provided awards and recognition for the top-ranking pen of steers.
Results and Discussion
Cattle consigned to the Dakota Feeder Calf Show feedout project averaged 604.88 pounds upon delivery to the Carrington Research Extension Center Livestock Unit on Oct. 20, 2018. After an average 212-day feeding period, cattle averaged 1,325.4 pounds (at plant, shrunk weight). Death loss was 2.33 percent (three head) during the feeding period.
Average daily feed intake per head was 36.2 pounds on an as-fed basis and 23.9 pounds on a dry-matter basis. Pounds of feed required per pound of gain were 11.1 on an as-fed basis and 7.35 pounds on a dry-matter basis.
The overall feed cost per pound of gain was $0.487. The overall yardage cost per pound of gain was $0.106. The combined cost per pound of gain, including feed, yardage, veterinary, trucking and other expenses except interest, was $0.746.
Calves were priced by weight upon delivery to the feedlot. The pricing equation ($ per 100 pounds = (-0.058517799* initial calf weight, pounds) + 198.6623157) was determined by regression analysis on local livestock auction prices reported for the weeks before and after delivery.
Overall, the carcasses contained U.S. Department of Agriculture Quality Grades at 2.4% Prime, 67.2% Choice (including 13.6% Certified Angus Beef), 28% Select, 0.8% Standard and 1.6% other, and USDA Yield Grades at 9.6% YG1, 44% YG2, 41.6% YG3, 4.8% YG4 and 0% YG5.
Carcass value per 100 pounds (hundredweight, or cwt) was calculated using the actual base carcass price plus premiums and discounts for each carcass. The grid price received for May 22, 2019, was $189.05 Choice YG3 base with premiums: Prime $15, CAB $6, YG1 $6.50 and YG2 $3, and discounts: Select minus $13, Standard (no roll) minus $15, YG4 minus $6 and defect carcasses (blood splash or dark cutter) minus $55.
Results from the calves selected for the pen-of-three competition are listed in Table 1.
Table 1. Feeding performance - 2018-2019 Dakota Feeder Calf Show Feedout
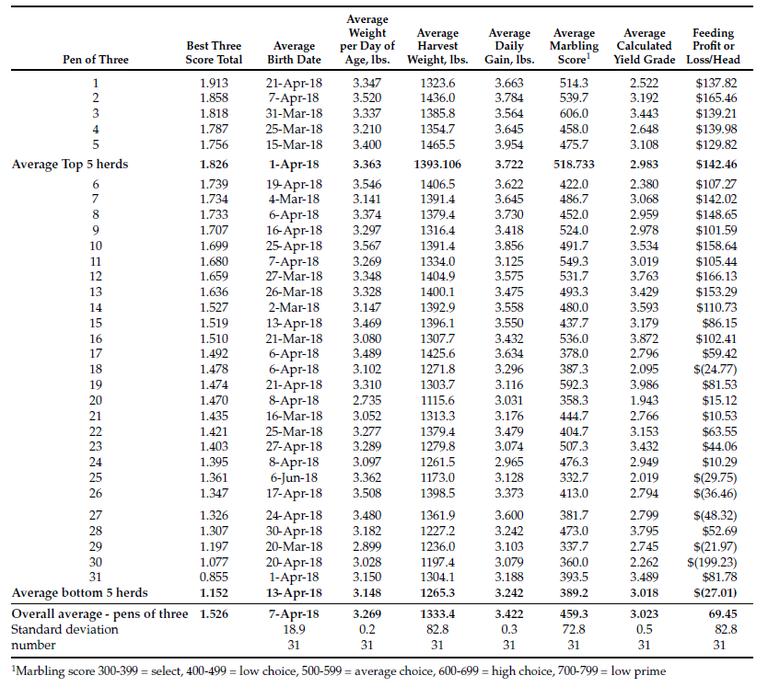
Table 2. Correlation between profit and various production measures (pen-of-three).
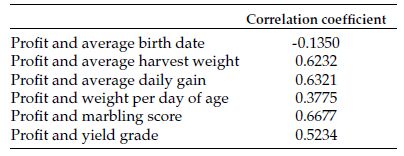
Overall, the pen-of-three calves averaged 409 days of age and 1,333.3 pounds per head at harvest. The overall pen-of-three finishing average daily gain was 3.42 pounds, while weight gain per day of age was 3.27 pounds. The overall pen-of-three marbling score was 459.3 (low choice, small marbling).
Correlations between profit and average birth date, harvest weight, average daily gain, weight per day of age or marbling score are shown in Table 2. Average harvest weight, average daily gain and marbling score had higher correlations to profitability than average birth date, average weight per day of age or yield grade.
The top-profit pen-of-three calves with superior genetics returned $167.46 per head, while the bottom pen-of-three calves returned minus $199.23 per head. The average of the five top-scoring pens of steers averaged $142.46 per head, while the average of the bottom five scoring pens of steers averaged minus $27.01 per head.
For the pen-of-three competition, average profit was $69.45 per head. The spread in profitability between the top and bottom five herds was $169.47 per head.
Implications
Calf value is improved with superior carcass and finishing performance. Exceptional average daily gains, weight per day of age, harvest weight and marbling score can be found in North Dakota beef herds. Feedout projects provide a source of information for cattle producers to learn about finishing performance and individual animal differences, and discover cattle value.
Literature Cited
Hoppe, K.F., V.L. Anderson, H. Hughes and K. Alderin. 1997. Finishing North Dakota Calves in North Dakota or Kansas - Final Report. A Report on Agricultural Research and Extension in Central North Dakota. 38:7. 10.
Effects of pen cleaning on feedlot performance and carcass characteristics of beef steers fed during the winter in the northern Great Plains
Bryan W. Neville1 and Rebecca L. Moore1
1Carrington Research Extension Center, NDSU
The objective of this study was to determine the effects of winter pen cleaning methods on feedlot performance and carcass characteristics of steers. Pen cleaning treatments did not influence animal performance within the conditions of this study. However, marbling score and quality grade of carcasses tended to be greater with increasing amounts of pen cleaning. The relationship between pen space and pen cleaning requires further research to determine their combined influences on animal performance.
Summary
Our hypothesis was that pen cleaning during the winter would improve animal performance and carcass characteristics of steers fed in the northern Great Plains. This study utilized 156 steer calves. Calves were assigned randomly to one of 12 pens, with pen randomly assigned to treatment. Treatments consisted of 1) Control - no cleaning, 2) Full - entirety of the pen cleaned twice throughout the study and 3) Apron - bunk aprons (approximately 10 feet) behind the bunk cleaned twice throughout the study. Calf weights were collected twice, prior to the start of study and prior to slaughter. Calves were fed for 195 days prior to reaching market readiness. Following slaughter, carcass data were collected. Our results are contrary to our hypothesis because pen cleaning resulted in no differences (P ≥ 0.48) in body weight, average daily gain (ADG), dry-matter intake (DMI) or feed efficiency in the current study. Hot carcass weight, ribeye area, back fat and yield grade of carcass also were not affected (P ≥ 0.44; Table 2) by pen cleaning treatment. We observed a tendency (P = 0.09) for greater marbling score and quality grade in carcasses resulting from steers managed with more thorough pen cleaning strategies. That pen cleaning did not improve animal performance was unexpected. It is possible that the light stocking density of pens or the ample bedding supplied negated the anticipated differences.
Introduction
The purpose of this study was to evaluate the efficacy of pen cleaning as a method to improve livestock performance of beef cattle fed to finish during the winter months in the northern Great Plains. Previous research with bedding frequency (Anderson et al., 2007) has demonstrated factors related to environment can improve animal performance.
Observing greater performance with pen cleaning as a result of reduced energy expenditure to maintain body temperature or by decreasing the energy expenditure associated with movement is logical. Secondary benefits to removing manure, mud and snow across the entirety of a dry lot pen would be captured through decreased maintenance of pen surfaces.
The removal of manure buildup directly behind the bunk line, allowing cattle easier access to feed, is a less labor-intensive cleaning method. Both pen cleaning methods have positive attributes, but no research has directly compared the extent of pen cleaning (cleaning bunk apron only vs. cleaning full pens) on animal performance. The objective of this project was to evaluate pen cleaning method on feedlot performance and carcass characteristics of steers fed in a dry lot during the winter in the northern Great Plains.
Experimental Procedures
This study was approved by the North Dakota State University Institutional Animal Care and Use Committee prior to initiation of study procedures. To accomplish our research objective, 156 beef steers (626.2 ± 30.28 pounds) were assigned to one of 12 pens (n = four per treatment).
Thirteen steers initially were placed into each pen. Pens were stocked at a similar density, approximately 290 square feet of pen space per animal. Treatment was assigned randomly to pen and consisted of: 1) Control - no cleaning, 2) Full - entirety of the pen cleaned twice throughout the study and 3) Apron - bunk aprons (approximately 10 feet) behind the bunk cleaned twice throughout the study.
Pens or aprons were cleaned on approximately 56-day intervals from the start of the study. Throughout the study, cattle were provided fresh bedding weekly, with an estimated 5.5 pounds/head/day of straw used during the course of the study.
Upon arrival, cattle were acclimated to pens. Weights were collected on two consecutive days and calves were sorted into pens. All calves received a growth promotant implant (Synovex S, Zoetis Inc., Parsippany-Troy Hills, N.J.) at the initiation of the study and were re-implanted 56 days later with Synovex Plus (Zoetis Inc., Parsippany-Troy Hills, N.J.).
Rations were developed to adapt cattle from a moderate-roughage diet to a high-concentrate diet. The final finishing ration consisted of 57.7% corn, 23.5% modified distillers grains with solubles (mDGS), 5% straw, 11% silage, 1.3% calcium carbonate and 1.5% supplement (dry-matter basis). Steers were fed for a total of 195 days. Feed was provided to target clean bunks the following morning prior to feeding.
At the conclusion of the feeding period, cattle were weighed on two consecutive days and shipped to a commercial abattoir for slaughter and subsequent carcass data collection. Data were analyzed with the mixed procedures of SAS (SAS Ins. Inc., Cary, N.C.). All data were analyzed, with pen serving as the experimental unit. Significance was declared at P ≤ 0.05.
Results and Discussion
We found no influence (P ≥ 0.48; Table 1) of pen cleaning on initial body weight (BW), final BW or ADG. Likewise, dry-matter intake and feed efficiency were similar (P ≥ 0.55) across treatment. Hot carcass weight, ribeye area, back fat and yield grade of carcass also were not affected (P ≥ 0.44; Table 2) by pen cleaning treatment. We observed a tendency (P = 0.09) for greater marbling score and quality grade in carcasses resulting from steers managed with more thorough pen cleaning strategies.
The data presented in the current study are largely contrary to our hypothesis. The reason for the lack of feedlot performance differences in steers could be attributed to lack of separation of pen conditions, to the relatively light stocking density of pens or to the adequate bedding provided to all pens. Any of these factors could have resulted in pen conditions that negated the possible impacts of the treatments imposed.
Previous research at the Carrington Research Extension Center has demonstrated that pen management can influence animal performance. Average daily gains of steers were improved by 0.86 pound/day by providing bedding, when compared with cattle not provided bedding (Anderson et al., 2007). This improvement is likely due to reduced energy expenditure to maintain body temperature.
Previous research evaluating mud depth, bedding and temperature has demonstrated significant effects to livestock performance and economic returns (Mader, 2011). Cleaning frequency has been evaluated (Wilson et al., 2004); unfortunately this research was not able to determine effects on animal performance. Pen maintenance, especially pen cleaning, may decrease the energy expenditures associated with movement, reduce environmental stress, and subsequently improve overall gain and performance.
Table 1. Impacts of pen cleaning on feedlot performance of steers fed during the winter in the northern Great Plains.
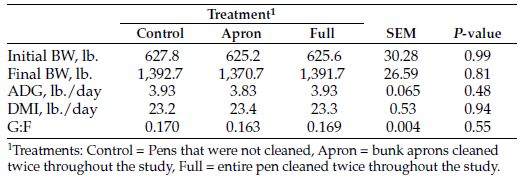
Table 2. Impacts of pen cleaning on carcass characteristics of steers fed during the winter in the northern Great Plains.
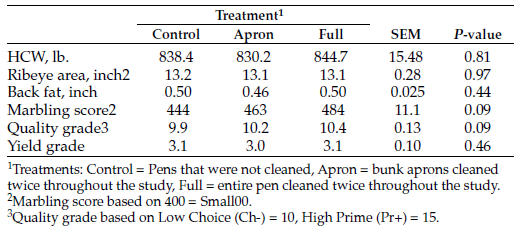
That pen cleaning did not improve animal performance in this study was unexpected. It is possible that the light stocking density of pens or the ample bedding supplied negated the anticipated differences.
Future research evaluating stocking density of pens and cleaning regimens may lead to better understanding of the benefits of pen cleaning. Specifically, research evaluating at what point animal space dictates that producers place greater attention on pen cleaning to avoid decreases in performance would be helpful for livestock producers. Additionally, continued research through time would prove beneficial because the differences in weather within a given year can influence cattle performance strongly.
Literature Cited
Anderson, V.L., and R.J. Wiederholt. 2007. The performance, economic, and environmental value of bedding feedlot cattle during northern Plains winters. J. Anim. Sci. 85:19. (Abstract)
Mader, T.L. 2011. Mud effects on feedlot cattle. 2011-Nebraska Beef Cattle Report. 82-83.
Wilson, C.B., G.E. Erickson, C.N. Macken and T.J. Klopfenstein. 2004. Impact of cleaning frequency on nitrogen balance in open feedlot pens. 2004-Nebraska Beef Cattle Report. 72-73.

(Photo by Rebecca Moore, NDSU)
Effects of a vitamin and mineral bolus on beef heifer feedlot performance, feeding behavior, carcass characteristics and liver mineral concentrations
Kacie L. McCarthy1, Sarah R. Underdahl1 and Carl R. Dahlen1
1Department of Animal Sciences, NDSU
The objectives of this study were to evaluate the effects of a vitamin and mineral bolus labelled for a 250-day duration on heifer feedlot performance, feeding behavior, carcass characteristics and liver mineral concentrations. Final body weight (BW), average daily gain (ADG), dry-matter intake (DMI), gain-to-feed (G:F) ratio and carcass characteristics were not influenced by bolus treatment. In this experiment, the vitamin and mineral bolus evaluated failed to increase liver mineral concentrations during the finishing period or influence heifer performance and carcass characteristics.
Summary
Crossbred beef heifers (n = 23; initial BW= 822 ± 25.6 pounds) housed at the Beef Cattle Research Complex (BCRC) in Fargo, N.D., were used to evaluate the effects of a vitamin and mineral bolus labelled for a 250-day duration on feedlot performance, feeding behavior, carcass characteristics and liver mineral concentrations. Heifers were assigned to one of two treatments: 1) received no supplemental mineral or vitamin (CON, n = 12) or 2) received two mineral and vitamin boluses on day zero (calcium [Ca], magnesium [Mg], sodium [Na], copper [Cu], iodine [I], manganese [Mn], selenium [Se], zinc [Zn], cobalt [Co|, vitamin A, vitamin D3 and vitamin E; Reloader 250, Cargill Inc., Minneapolis, Minn.; MIN, n = 11). Heifers were fed a total mixed ration (TMR) (16.05% crude protein [CP], 1.44 megacalories per kilogram of net energy gain [Mcal/kg NEg]) with no added vitamin or mineral supplement. Feed intake, and number and time of visits were recorded for each heifer using the Insentec feeding system (Hokofarm Group B.V., the Netherlands) during the 150-day feeding period. Liver biopsies were collected from heifers on days zero, 69 and 134 of the feeding period for analysis of mineral concentrations. Heifers were slaughtered after 150 days on feed and carcass characteristics were determined. Final BW, ADG, DMI, G:F and carcass characteristics were not influenced (P > 0.19) by treatment. Control heifers visited feeders more but spent less time per visit and ate less per visit, compared with MIN heifers (P < 0.03). No differences (P > 0.06) in liver mineral concentrations were observed between treatments. Liver mineral concentrations for both treatments were decreasing through time for Se, Cu, molybdenum (Mo), Mn and Co (P < 0.05). In this experiment, the slow-release vitamin and mineral bolus evaluated failed to increase liver mineral concentrations during the finishing period or influence heifer performance and carcass characteristics.
Introduction
Diet alone often does not supply sufficient amounts of trace minerals; therefore, supplementation is necessary to optimize animal health and performance (NASEM, 2016). Research clearly has documented that intakes of free-choice minerals are variable among animals, with some cattle over or underconsuming supplements (Greene, 2000). Furthermore, inadequate trace mineral consumption can compromise reproduction, animal health and animal growth (NRC, 2005; NASEM, 2016).
A long-acting (six-month) reticulorumen trace mineral bolus containing Cu, Se and Co has been developed in the United Kingdom (Cosecure; Telsol Ltd., Leeds, UK) and has shown promise in increasing liver copper and blood selenium levels in range cows (Sprinkle et al., 2006). A recently developed bolus product has been released in the U.S. (Reloader 250, Cargill Inc., Minneapolis, Minn.) that targets a slow release for 250 days. A long-acting delivery of trace minerals could be advantageous for beef production in extensively managed systems.
In confinement scenarios, minerals and vitamins usually are provided as loose or pelleted supplements that are balanced properly in the diet. However, in range or pasture settings, range pellets can be delivered to cattle, but the inconvenience of transporting large quantities of feed to remote grazing sites often has producers providing mineral and protein supplements on a free-choice basis. This method assumes that cattle will consume the quantities that are recommended and satisfy requirements.
Therefore, this study was a model to evaluate the slow-release bolus in a confinement scenario prior to testing the bolus in extensive environments. The objectives of this study were to evaluate effects of a vitamin and mineral bolus on heifer feedlot performance, feeding behavior, carcass characteristics and liver mineral concentrations.
Experimental Procedures
All animal procedures were conducted in accordance with the Institutional Animal Care and Use Committee at North Dakota State University.
Animal Procedures
Twenty-three crossbred beef heifers (initial BW= 822 ± 25.6 pounds) were housed at the Beef Cattle Research Complex (BCRC) in Fargo, N.D. Heifers were assigned to one of two treatments: 1) received no supplemental mineral or vitamin bolus (CON, n = 12), or 2) received two mineral and vitamin boluses on day zero (Ca, Mg, Na, Cu, I, Mn, Se, Zn, Co, vitamin A, vitamin D3 and vitamin E; Reloader 250, Cargill Inc., Minneapolis, Minn.; MIN, n = 11). Heifers were fed a TMR that consisted of corn silage, grass hay, corn and dried distillers grain plus solubles (Table 1; 1.44 Mcal/kg NEg) with no added vitamin or mineral supplement.
Feed intake for each heifer was monitored using the Insentec feeding system (Hokofarm Group B.V., the Netherlands) during the 150-day feeding period. Feed intake and behavior were summarized by day for each individual heifer (Montanholi et al., 2010; Swanson et al., 2014).
Liver biopsies were collected from heifers on days zero, 69, and 134 of the feeding period to measure mineral concentrations. Liver samples were sent to Michigan State University and were evaluated for concentrations of minerals using inductively coupled plasma mass spectrometry.
Carcass characteristics were collected at slaughter at a commercial abattoir. Hot carcass weight data were obtained following animal slaughter, whereas marbling score, back fat, longissimus muscle area and yield grade were taken after carcasses were chilled for 24 hours.
Table 1. Dietary ingredient and nutrient composition.
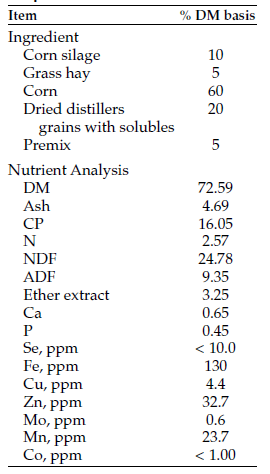
Statistical Analysis
Data were analyzed using the GLM procedure of SAS (9.4, SAS Inst. Inc., Cary, N.C.) for intake, behavior and carcass characteristics. Liver mineral concentrations were analyzed using the MIXED procedure as repeated measures with the model statement containing the effects of treatment, time and the interaction of treatment and time.
The covariance structure used was variance components. For all analysis, individual animal was the experimental unit and results are reported as least square means with significance at P < 0.05.
Results and Discussion
Final body weight did not differ between treatments (Table 2; P = 0.77). We found no differences (P = 0.93) in ADG between treatments, with heifers gaining 2.75 pounds per day. No differences (P = 0.23) were observed in dry-matter intake between treatments, with heifers consuming 21.6 pounds per day.
Feed efficiency (G:F) also was not influenced (P = 0.41) by treatment and averaged 0.283 pounds per pound. In similar studies, performance responses have been variable relative to trace mineral or Cu and Zn supplementation on finishing steer performance with no effect on ADG, DMI or G:F (Spears and Kegley, 2002; Ahola et al., 2005) versus nonsupplemented controls; whereas DMI has been shown to decrease as Zn supplementation has increased (Malcolm-Callis et al., 2000).
Control heifers visited the feeders more frequently (P = 0.01), compared with heifers receiving the mineral bolus. However, the number of meals per day was not influenced (P = 0.96) by treatment.
Heifers receiving the mineral bolus ate longer (P = 0.01) per visit than control heifers. However, the amount of time heifers spent eating per meal and per day was not different (P > 0.49) between treatments. Heifers receiving the mineral bolus ate more per visit (P = 0.03), compared with control heifers, but we found no differences (P > 0.08) in eating rate per meal and eating rate per minute between treatments.
No differences (Table 3; P = 0.86) were observed between treatments for hot carcass weight, which averaged 61%. Modest marbling ranges from 500 to 600 (Emerson et al., 2013); moreover, heifers in the current report fall within the range of 584 and 523 for CON and MIN heifers, respectively.
Table 2. Feed intake and feeding behavior of heifers receiving vitamin and mineral bolus.
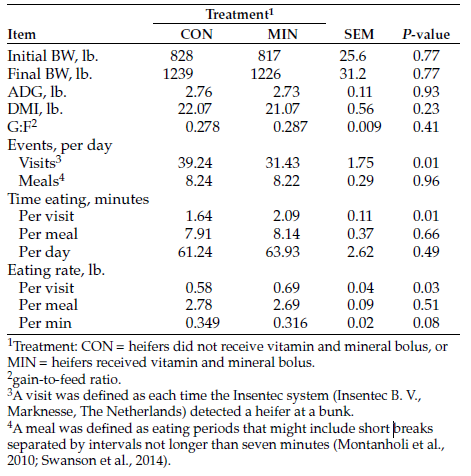
Table 3. Carcass characteristics from heifers receiving vitamin and mineral bolus.
We found no differences (P > 0.38) in backfat, longissimus area and yield grade between treatments. Reports of zinc supplementation on finishing phase carcass characteristics have found zinc to increase quality grade and marbling in steers, compared with nonsupplemented steers (Spears and Kegley, 2002).
Liver mineral concentrations were not different between treatments (P > 0.06) for Se, iron (Fe), Cu, Zn, Mn, Mo and Co (Table 4). However, liver mineral concentrations decreased through time for Se, Cu, Mo, Mn and Co (P < 0.05) and increased (P < 0.001) through time for Fe.
By the end of the period, Cu liver concentrations would be considered marginal (33 to 125 micrograms per gram [μg/g] DM; Kincaid, 2000). As defined by Kincaid (2000), the heifers in this study were marginally deficient (7 to 15 micrograms per gram [μg/g] dry matter [DM]) for Mn.
Table 4. Liver mineral concentrations from heifers receiving vitamin and mineral bolus.
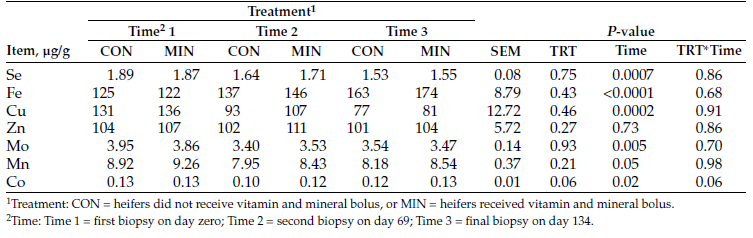
Heifer liver Se concentrations fall within adequate ranges (1.25 to 2.50 micrograms per gram [μg/g] dry matter [DM]; Kincaid, 2000). Likewise, liver Co concentrations fall within the satisfactory range of 0.08-0.12 μg/g DM (McNaught, 1948).
In this experiment, the slow-release vitamin and mineral bolus evaluated failed to increase liver mineral concentrations during the finishing period or influence heifer performance and carcass characteristics.
Acknowledgments
The authors acknowledge the staff at the Beef Cattle Research Complex for assistance with animal handling and feeding.
Literature Cited
Ahola, J.K., L.R. Sharpe, K.L. Dorton, P.D. Burns, T.L. Stanton and T.E. Engle. 2005. Effects of Lifetime Copper, Zinc, and Manganese Supplementation and Source on Performance, Mineral Status, Immunity, and Carcass Characteristics of Feedlot Cattle. Prof. Anim. Sci. 21:305-317.
Emerson, M.R., D.R. Woerner, K.E. Belk and J.D. Tatum. 2013. Effectiveness of USDA instrument-based marbling measurements for categorizing beef carcasses according to differences in longissimus muscle sensory attributes1. J. Anim. Sci. 91:1024-1034.
Greene, L.W. 2000. Designing mineral supplementation of forage programs for beef cattle. J Anim Sci 77:1-9.
Kincaid, R.L. 2000. Assessment of trace mineral status of ruminants: A review. J. Anim. Sci. 77(Suppl E):1-10.
Malcolm-Callis, K.J., G.C. Duff, S.A. Gunter, E.B. Kegley and D.A. Vermeire. 2000. Effects of supplemental zinc concentration and source on performance, carcass characteristics, and serum values in finishing beef steers. J. Anim. Sci. 78:2801-2808.
McNaught, K.J. 1948. Cobalt, copper and iron in the liver in relation to cobalt deficiency ailment. New Zealand Journal of Science and Technology, Section A 30:26-43.
Montanholi, Y.R., K.C. Swanson, R. Palme, F.S. Schenkel, B.W. McBride, D. Lu and S.P. Miller. 2010. Assessing feed efficiency in beef steers through feeding behavior, infrared thermography and glucocorticoids. Animal 4:692-701.
NASEM. 2016. Nutrient requirements of beef cattle: Eighth revised edition. National Academies Press, Washington, D.C.
NRC. 2005. Mineral tolerance of animals. Second Revised Edition ed. The National Academies Press.
Spears, J.W., and E.B. Kegley. 2002. Effect of zinc source (zinc oxide vs. zinc proteinate) and level on performance, carcass characteristics, and immune response of growing and finishing steers. J. Anim. Sci. 80:2747-2752.
Sprinkle, J.E., S.P. Cuneo, H.M. Frederick, R.M. Enns, D.W. Schafer, G.E. Carstens, S.B. Daugherty, T.H. Noon, B.M. Rickert and C. Reggiardo. 2006. Effects of a long-acting, trace mineral, reticulorumen bolus on range cow productivity and trace mineral profiles. J. Anim. Sci. 84:1439-1453.
Swanson, K.C., A. Islas, Z.E. Carlson, R.S. Goulart, T.C. Gilbery and M.L. Bauer. 2014. Influence of dry-rolled corn processing and increasing dried corn distillers grains plus solubles inclusion for finishing cattle on growth performance and feeding behavior. J. Anim. Sci. 92:2531-2537.
Ractopamine hydrochloride supplementation influences feeding behavior, growth performance and carcass characteristics of finishing steers
Ronald J. Trotta1, Kasey R. Maddock-Carlin1 and Kendall C. Swanson1
1Department of Animal Sciences, NDSU
The objectives of this experiment were to evaluate the effects of feeding ractopamine hydrochloride (RAC) for 42 days to finishing steers on feeding behavior, growth performance and carcass characteristics. Data from the current study demonstrated that RAC supplementation for 42 days improves feed efficiency, increases the rate of dry-matter intake (DMI) without altering DMI, and increases muscling in finishing cattle.
Summary
Ractopamine hydrochloride (RAC) is a beta-adrenergic agonist that functions as a repartitioning agent to improve muscling in finishing cattle. Many studies have investigated the effects of RAC on growth performance and carcass characteristics; however, minimal information is available about the influence of RAC on feeding behavior. Sixty-nine steers [body weight (BW) = 802 ± 8.6 pounds] predominately of Angus and Simmental breeding were subjected to a 126-day (n = 46) or 154-day (n = 23) feeding period and assigned randomly to one of two treatment groups: supplementation to provide 0 (CON; n = 34) or 267 ± 4.9 milligrams per day (mg/d) of RAC (n = 35). Ractopamine was provided as Optaflexx 45 at 0.024% of the diet (dry matter [DM] basis; Elanco Animal Health, Greenfield, Ind.). Dietary treatments were fed the final 42 days in the feed yard (treatment period). Feeding behavior and growth performance were measured using radio frequency identification tags and the Insentec feeding system. Following the final day of treatment, steers were slaughtered and carcass measurements were recorded. Data were analyzed using MIXED models in SAS. We found no differences in BW, average daily gain (ADG), DMI, gain:feed (G:F) or feeding behavior during the pretreatment period (P > 0.44). Ractopamine supplementation increased G:F during the treatment period (P = 0.02) and during the total period (P = 0.03), and tended to increase ADG during the treatment and total period (P ≤ 0.08). Dry matter intake was not affected during the treatment or total period (P > 0.67). Eating time per visit, per meal and per day were decreased (P < 0.02) in steers supplemented with RAC during the treatment period. Dry matter intake per minute was increased (P = 0.02) in steers supplemented with RAC. Hot carcass weight, dressing percentage and 12th rib fat were not influenced by RAC supplementation. Ractopamine supplementation decreased marbling (P = 0.008) and kidney, pelvic and heart percentage (P = 0.04), and increased longissimus muscle area (P = 0.01). These data demonstrate that RAC supplementation for 42 days improves feed efficiency, increases the rate of DMI without altering DMI, and increases muscling in finishing cattle.
Introduction
Dietary supplementation of beta-adrenergic receptor agonists have shown improvements in growth performance and carcass characteristics of various livestock species, including cattle, sheep, poultry and swine. Ractopamine hydrochloride (RAC) is the most commonly used beta-adrenergic receptor agonist in the U.S. for finishing beef cattle production (Samuelson et al., 2016). Functioning as a repartitioning agent (Moloney et al., 1991), RAC increases average daily gain (ADG), hot carcass weight (HCW) and longissimus muscle area (LMA) with minimal effects on adipose tissue (Lean et al., 2014).
We found multiple reports that RAC supplementation can increase ADG and gain:feed (G:F) in finishing cattle during the last 28 to 42 days on feed without changing dry matter intake (DMI). However, information on how RAC affects feeding behavior of finishing cattle is limited. Moreover, no reports are available about the effects of feeding RAC during a longer period (42 days) on feeding behavior in steers.
Increasing the length of feeding of RAC in a complete feed is recommended for increased ADG, G:F and carcass leanness. Therefore, the objectives of this experiment were to evaluate the effects of feeding RAC for 42 days to finishing steers on feeding behavior, growth performance and carcass characteristics.
Experimental Procedures
All animal procedures were approved by the North Dakota State University Institutional Animal Care and Use Committee.
Sixty-nine steers [body weight (BW) = 802 ± 8.6 pounds predominately of Angus and Simmental breeding were stratified by weight into three pens (n = 23 per pen) and were subjected to a 126- (n = 46) or 154-day (n = 23) feeding period and assigned randomly to dietary treatment group within pen. The total feeding period consisted of the pretreatment and treatment periods.
The treatment period was a fixed length of 42 days. Thus, the pretreatment period was either 84 or 112 days. Steers were fed a common concentrate-based finishing diet containing monensin (Elanco Animal Health, Greenfield, Ind.) for all periods in the trial. The total mixed rations were delivered once daily at 8 a.m. with a feed delivery truck. Steers had ad libitum access to feed and water.
On the final day of the pretreatment period, steers were weighed (BW = 1,184 ± 14.1 pounds) on two consecutive days and began the treatment period: supplementation to provide 0 milligrams per day (mg/d) RAC (CON; n = 34) or RAC (267 ± 4.9 mg/d; n = 35) for the last 42 days in the feed yard. For the RAC treatment, Optaflexx 45 (Elanco Animal Health, Greenfield, Ind.) was mixed into the diet at 0.024% of dry matter (DM) as part of the supplement (5% of DM).
Radio frequency identification tags were placed in the right ear of each steer prior to the beginning of the experiment. Each pen contained eight automated electronic feeding stations (Insentec, Hokofarm Group, Marknesse, The Netherlands; four feeding stations dedicated for each treatment) as described previously (Swanson et al., 2014), allowing for offering specific dietary treatments and monitoring of individual feed intake and feeding behavior characteristics.
Following the 42-day treatment period, steers were slaughtered at a commercial packing facility and carcass traits were measured. All data were analyzed using the MIXED procedure of SAS.
Results and Discussion
We observed no differences in growth performance and feeding behavior traits in the pretreatment period (data not shown). Therefore, no differences occurred between groups before the initiation of the RAC treatment period that could have influenced treatment responses.
Ractopamine supplementation increased G:F during the treatment period (P = 0.02) and during the total period (P = 0.03), and tended to increase ADG during the treatment and total period (P ≤ 0.08). The observed improvement in feed efficiency (G:F) with RAC supplementation was likely because of its function as a repartitioning agent (Moloney et al., 1991).
Hot carcass weight, dressing percentage and 12th rib fat were not influenced by RAC supplementation. Ractopamine supplementation decreased marbling (P = 0.008) and kidney, pelvic and heart percentage (P = 0.04), and increased longissimus muscle area (P = 0.01). The reductions in KPH and marbling score and increases in LMA in this study are carcass indicators for effects of repartitioning of energy from adipose tissue by RAC.
Table 1. Influence of ractopamine hydrochloride supplementation to finishing steers on feeding behavior.1
Table 2. Influence of ractopamine hydrochloride supplementation to finishing steers on growth performance and carcass characteristics.1
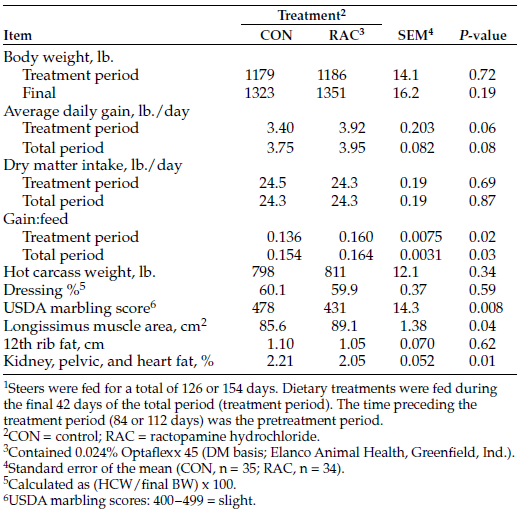
Dry matter intake was not affected during the treatment or total period (P > 0.67). Eating time per visit, per meal and per day were decreased (P < 0.02) in steers supplemented with RAC during the treatment period. Dry matter intake per minute was increased (P = 0.02) in steers supplemented with RAC. This increase in eating rate (13%) for steers supplemented with RAC was likely the driving factor for reducing time eating per day (13.7%), compared with CON steers.
In conclusion, feeding ractopamine hydrochloride at 267 mg/d for 42 days influenced feeding behavior, growth performance and carcass characteristics of finishing steers. Ractopamine hydrochloride supplementation tended to increase ADG and improve feed efficiency.
Evidence of repartitioning was observed through reductions in marbling score and KPH percentage, and an increase in LMA. Supplementation of ractopamine increased DMI per minute and subsequently reduced eating time by 13% per day.
Although the biological significance of the effects of RAC on rate of DMI are not clear, we note potential concerns related to feeding management to avoid digestive disturbances, such as ruminal acidosis. Future research should be directed toward investigating effects of RAC supplementation on feeding behavior and associated potential for influencing rumen fermentation and gastrointestinal function.
Acknowledgments
This work was partially supported by the North Dakota Beef Commission.
Literature Cited
Lean, I.J., J.M. Thompson and F.R. Dunshea. 2014. A meta-analysis of zilpaterol and ractopamine effects on feedlot performance, carcass traits and shear strength of meat in cattle. PLoS ONE. 9:e115904.
Moloney, A., P. Allen, R. Joseph and V. Tarrant. 1991. Influence of beta-adrenergic agonists and similar compounds on growth. In: Growth Regulation in Farm Animals. Advances in Meat Research, vol. 7. pp 455-513. Elsevier Applied Science, New York, NY.
Samuelson, K.L., M.E. Hubbert, M.L. Galyean and C.A. Löest. 2016. Nutritional recommendations of feedlot consulting nutritionists: The 2015 New Mexico State and Texas Tech University survey. J. Anim. Sci. 94:2648–2663.
Swanson, K.C., A. Islas, Z.E. Carlson, R.S. Goulart, T.C. Gilbery and M.L. Bauer. 2014. Influence of dry-rolled corn processing and increasing dried corn distillers grains plus solubles inclusion for finishing cattle on growth performance and feeding behavior. J. Anim. Sci. 92:2531– 2537.
Meat Science
Tenderness and juiciness of beef steaks from varying hot carcass weights
Michaella Fevold1, Laura Grube1, Wanda Keller1, Kasey Maddock-Carlin1, Robert Maddock1
1Department of Animal Sciences, NDSU
The objective of this study was to determine how hot carcass weights affect temperature decline and pH decline of carcasses, as well as, tenderness and juiciness of steaks. Hot carcass weight did not have an influence on overall meat quality attributes of steaks.
Summary
Beef carcasses were separated into light, medium or heavy weight groups and temperature and pH decline were measured for 24 hours. Tenderness and juiciness were measured objectively on steaks from each carcass. Hot carcass weight did not have an influence on overall meat quality attributes of steaks.
Introduction
Tenderness is a main driver in beef consumer satisfaction and encourages return customers (Boleman et al., 1997). With this knowledge, research needs to be conducted continually in the U. S. to better understand what affects beef tenderness. In 2015, the U. S. produced more beef than in 1977 with 13 million fewer cattle harvested (Maples et al., 2018).
This increase can be attributed to many improved production methods, including better genetics and nutrition. Increasing the efficiency of beef production in the U. S. has caused live cattle and carcass weights to increase with an increase in carcass weight of almost 100 pounds in the past decade (Maples et al., 2018). In 2016, the National Beef Quality Audit reported that carcass weight and size were considered a top six priority area, behind food safety, eating satisfaction and lean fat and bone (NBQA, 2016).
The objectives of this research were to evaluate chilling rate and pH decline of carcasses from different weights classes and how they relate to tenderness and juiciness of longissimus and semimembranosus steaks.
Experimental Procedures
Beef carcasses (n = 59) were selected at a commercial abattoir (DemKota Ranch Beef, Aberdeen, S.D.). Carcasses were selected after completion of the harvest process and before moving them into chill coolers. Carcasses were selected in various weight ranges: light (less than 800 pounds), medium (800 to 900 pounds), and heavy (greater than 900 pounds).
Muscle pH was measured immediately from the center of the semimembranosus and the longissimus muscle between the 12th and 13th rib. Muscle pH measurements were taken in the same location at four and 24 hours after initial measurements. Additionally, temperature was recorded for 24 hours using a thermometer probe inserted into the longissimus and semimembranosus muscle.
After carcasses were chilled for approximately 24 hours, carcass data including ribeye area (REA), 12th rib backfat (BF), kidney, pelvic and heart fat percentage (KPH), marbling score and U.S. Department of Agriculture final yield grade were collected. Upon fabrication, ribeye rolls (IMPS 112a) and inside rounds (IMPS 160a) were transferred to the North Dakota State University Meat Laboratory in vacuum-sealed bags.
After subprimals were aged for 14 days, three test steaks were fabricated from each ribeye roll and two tests steaks were fabricated from each inside round. Drip loss, cooking loss, and Warner-Bratzler shear force (WBSF) were taken from each steak.
Results and Discussion
Temperature decline
No differences were observed among hot carcass weight groups in longissimus muscle temperature at zero hours and 24 hours (Table 1). In addition, no differences were observed among hot carcass weight groups in semimembranosus muscle temperature at zero hour and four hours. Longissimus muscle temperature at four hours was lower in carcasses classified as light weight compared with carcasses classified as heavy weight. Additionally, semimembranosus muscle temperature at 24 hours was lower in carcasses classified as light and medium weight compared with carcasses classified as heavy weight.
Table 1. Least squares means ± standard error of means of the relationship among hot carcass weight and temperature decline in degrees Fahrenheit of beef longissimus and semimembranosus muscle.
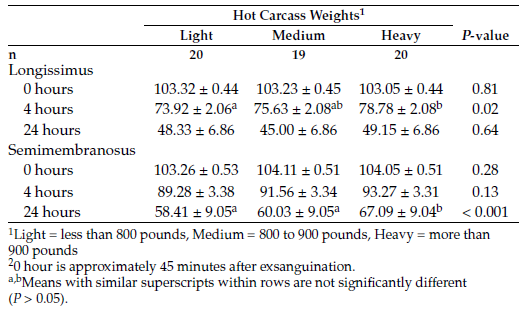
Table 2. Least squares means of the relationship among hot carcass weight and beef carcass characteristics.
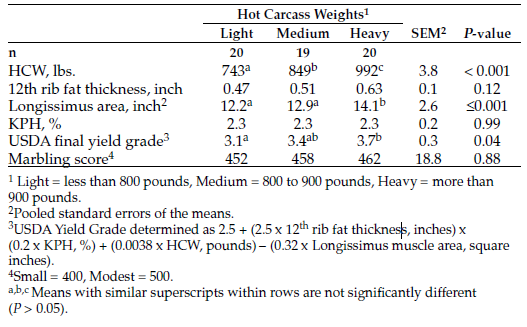
These differences are in agreement with several papers (Lochner et al., 1979; Jones and Robertson, 1988; Okeudo and Moss, 2005) which observe a difference in muscle temperature decline influenced by fat cover. While 12th rib fat thickness was not statistically different, we observed differences in fat thickness between light and heavy weight carcasses.
Muscle pH decline
No differences among hot carcass weight groups were observed for longissimus or semimebranosus muscle pH at zero hour, four and 24 hours. These results were expected because glycogen levels would not be different based on hot carcass weight and therefore would not affect muscle pH decline (Pethick et al., 1995).
Carcass characteristics
No differences were observed among hot carcass weight groups for fat thickness, kidney, pelvic and heart fat percentage and marbling score (Table 2). Longissimus area differed between carcasses classified with light and medium weight groups having smaller longissimus areas than those classified as heavy weight.
In addition, USDA final yield grade differed between carcasses with light weight carcasses having lower USDA yield grades, compared with carcasses classified as heavy weight. These results were expected due to the known relationship between increasing hot carcass weight and larger longissimus areas and higher USDA final yield grades (Nour et al., 1983).
Drip loss, tenderness and cook loss
No differences were observed in drip loss, WBSF and cook loss of longissimus and semimembranosus muscle among hot carcass weight groups. Results for drip loss and cook loss are consistent with the literature that the water-holding capacity of raw and cooked meat is the most compromised when pH falls below the normal range of 5.6 to 5.8 (Huff-Lonergan, 2010). In addition, results for tenderness also were in agreement with current literature, which indicates the rate of pH and temperature decline have a significant influence on beef tenderness (Lochner et al., 1979; Hannula and Puolanne, 2004).
Implications
Our results may have different implications to different segments of the beef industry and consumers of beef products. Beef producers should view our results with caution. While we did not see an influence on tenderness or juiciness of beef steaks due to increased hot carcass weights, producers do need to be aware research is showing that consumers discriminate against larger steaks because of the thickness of steaks, and continued increase in hot carcass weights could result in consumers switching to another protein source such as pork or chicken.
Beef producers also should take note that our research indicates fat cover is important in the chilling of beef carcasses and likely has some influence on producing a tender product for consumers. Beef packers also should take note of these observations and consider discouraging producers from sending underfinished cattle to be harvested to avoid an increase in toughness of beef products. Beef packers also may want to consider reaching out to their retail and restaurant customers to gather information on if cut size has been an issue to stay ahead of any potential problems.
Literature Cited
Boleman, S.J., Boleman, S.L., Miller, R.K., Taylor, J.F., Cross, H.R., Wheeler, T.L., Koohmaraie, M., Shackleford, S.D., Miller, M.F., West, R.L., Johnson, D.D., Savell, J.W. 1997. Consumer evaluation of beef of known categories of tenderness. J. Anim. Sci. 75: 1521-1524.
Hannula, T., and Puolanne, E. 2004. The effect of cooling rate on beef tenderness: the significance of pH at 7°C. Meat Sci. 67: 403-408.
Huff-Lonergan, E. 2010. Water holding capacity of fresh meat. National Pork Board and American Meat Science Association.
Jones, S.D.M., and Robertson, W.M. 1988. The effects of spray-chilling carcasses on the shrinkage and quality of beef. Meat Sci. 24: 177-188
Lochner, J.V., Kauffman, R.G., Marsh, B.B. 1979. Early postmortem cooling rate and beef tenderness. Meat Sci. 4: 227-241.
Maples, J.G., Lusk, J.L., Peel, D.S. 2018. Unintended consequences of the quest for increased efficiency in beef cattle: When bigger isn’t better. Food Policy. 74: 65-73
National Beef Quality Audit Executive Summary. 2016. National Cattleman’s Beef Association.
Nour, A.Y.M., Thonney, M.L., Stouffer, J.R., White, W.R.C. 1983. Changes in carcass weight and characteristics with increasing weight of large and small cattle. J. Anim. Sci. 57: 1154-1165.
Okeudo, N.J., and Moss, B.W. 2005. Interrelationships amongst carcass and meat quality characteristics of sheep. Meat Sci. 69: 1-8
Pethick, D.W., Rowe, J.B., Tudor, G. 1995. Glycogen metabolism and meat quality. Recent Advances in Animal Nutrition in Australia. 97-103.

(Photo by Kacie McCarthy, NDSU)
